

Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA
- PMID: 22344696
- PMCID: PMC3367185
- DOI: 10.1093/nar/gks144
Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA
Abstract
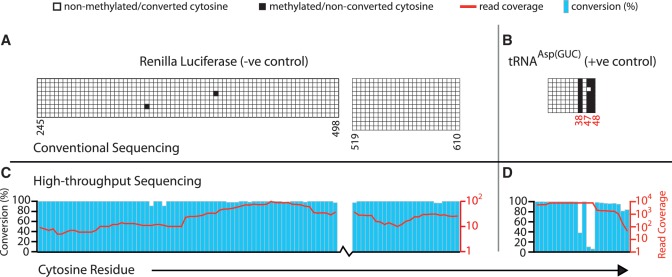
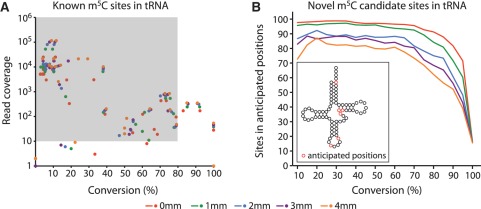
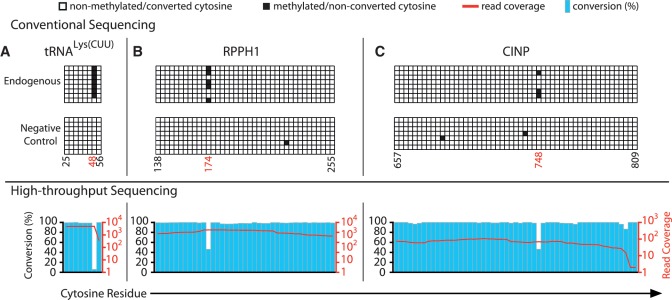
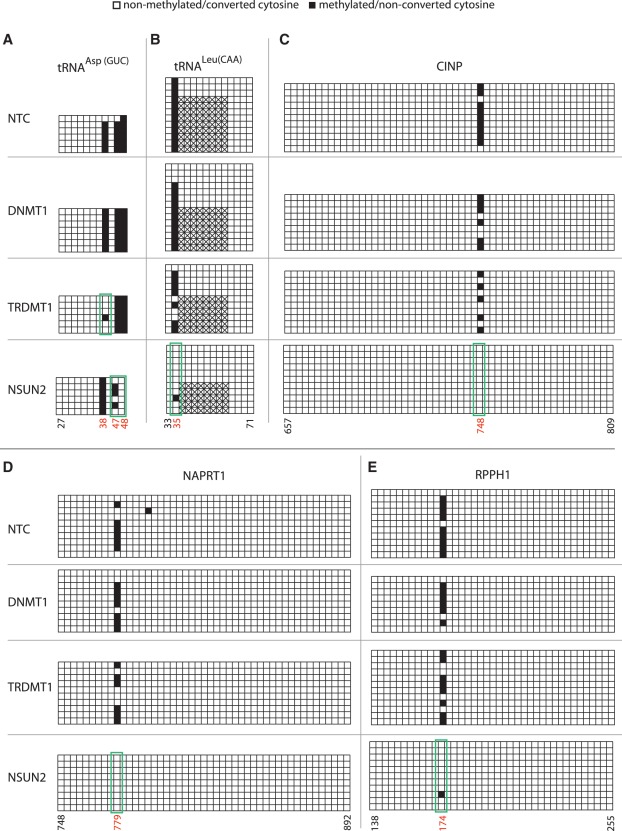
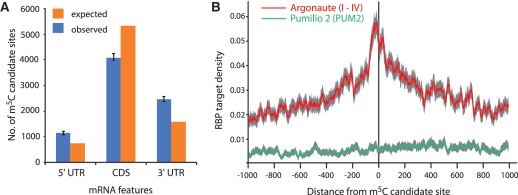
The modified base 5-methylcytosine (m(5)C) is well studied in DNA, but investigations of its prevalence in cellular RNA have been largely confined to tRNA and rRNA. In animals, the two m(5)C methyltransferases NSUN2 and TRDMT1 are known to modify specific tRNAs and have roles in the control of cell growth and differentiation. To map modified cytosine sites across a human transcriptome, we coupled bisulfite conversion of cellular RNA with next-generation sequencing. We confirmed 21 of the 28 previously known m(5)C sites in human tRNAs and identified 234 novel tRNA candidate sites, mostly in anticipated structural positions. Surprisingly, we discovered 10,275 sites in mRNAs and other non-coding RNAs. We observed that distribution of modified cytosines between RNA types was not random; within mRNAs they were enriched in the untranslated regions and near Argonaute binding regions. We also identified five new sites modified by NSUN2, broadening its known substrate range to another tRNA, the RPPH1 subunit of RNase P and two mRNAs. Our data demonstrates the widespread presence of modified cytosines throughout coding and non-coding sequences in a transcriptome, suggesting a broader role of this modification in the post-transcriptional control of cellular RNA function.
Figures
Similar articles
-
Conservation of tRNA and rRNA 5-methylcytosine in the kingdom Plantae.BMC Plant Biol. 2015 Aug 14;15:199. doi: 10.1186/s12870-015-0580-8. BMC Plant Biol. 2015. PMID: 26268215 Free PMC article.
-
Multiple links between 5-methylcytosine content of mRNA and translation.BMC Biol. 2020 Apr 15;18(1):40. doi: 10.1186/s12915-020-00769-5. BMC Biol. 2020. PMID: 32293435 Free PMC article.
-
Function and detection of 5-methylcytosine in eukaryotic RNA.Epigenomics. 2010 Oct;2(5):709-15. doi: 10.2217/epi.10.47. Epigenomics. 2010. PMID: 22122054
-
Eukaryotic 5-methylcytosine (m⁵C) RNA Methyltransferases: Mechanisms, Cellular Functions, and Links to Disease.Genes (Basel). 2019 Jan 30;10(2):102. doi: 10.3390/genes10020102. Genes (Basel). 2019. PMID: 30704115 Free PMC article. Review.
-
Post-transcriptional regulation by cytosine-5 methylation of RNA.Biochim Biophys Acta Gene Regul Mech. 2019 Mar;1862(3):240-252. doi: 10.1016/j.bbagrm.2018.12.003. Epub 2018 Dec 27. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 30593929 Review.
Cited by
-
Transcriptome-wide mapping of 5-methylcytidine RNA modifications in bacteria, archaea, and yeast reveals m5C within archaeal mRNAs.PLoS Genet. 2013 Jun;9(6):e1003602. doi: 10.1371/journal.pgen.1003602. Epub 2013 Jun 27. PLoS Genet. 2013. PMID: 23825970 Free PMC article.
-
Neuronal Depolarization Induced RNA m5C Methylation Changes in Mouse Cortical Neurons.Biology (Basel). 2022 Jun 29;11(7):988. doi: 10.3390/biology11070988. Biology (Basel). 2022. PMID: 36101370 Free PMC article.
-
Long non-coding RNAs as targets for cytosine methylation.RNA Biol. 2013 Jun;10(6):1003-8. doi: 10.4161/rna.24454. Epub 2013 Apr 1. RNA Biol. 2013. PMID: 23595112 Free PMC article.
-
Synthesis of 5-Hydroxymethylcytidine- and 5-Hydroxymethyl-uridine-Modified RNA.Synthesis (Stuttg). 2016 Apr;48(8):1108-1116. doi: 10.1055/s-0035-1561220. Synthesis (Stuttg). 2016. PMID: 27413246 Free PMC article.
-
Sex-Based Mhrt Methylation Chromatinizes MeCP2 in the Heart.iScience. 2019 Jul 26;17:288-301. doi: 10.1016/j.isci.2019.06.031. Epub 2019 Jun 27. iScience. 2019. PMID: 31323475 Free PMC article.
References
-
- Hotchkiss RD. The quantitative separation of purines, pyrimidines, and nucleosides by paper chromatography. J. Biol. Chem. 1948;175:315–332. - PubMed
-
- Suzuki MM, Bird A. DNA methylation landscapes: provocative insights from epigenomics. Nat. Rev. Genet. 2008;9:465–476. - PubMed
-
- Wyatt GR. Occurrence of 5-methylcytosine in nucleic acids. Nature. 1950;166:237–238. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
