

Enhancement of chemokine function as an immunomodulatory strategy employed by human herpesviruses
- PMID: 22319442
- PMCID: PMC3271085
- DOI: 10.1371/journal.ppat.1002497
Enhancement of chemokine function as an immunomodulatory strategy employed by human herpesviruses
Abstract
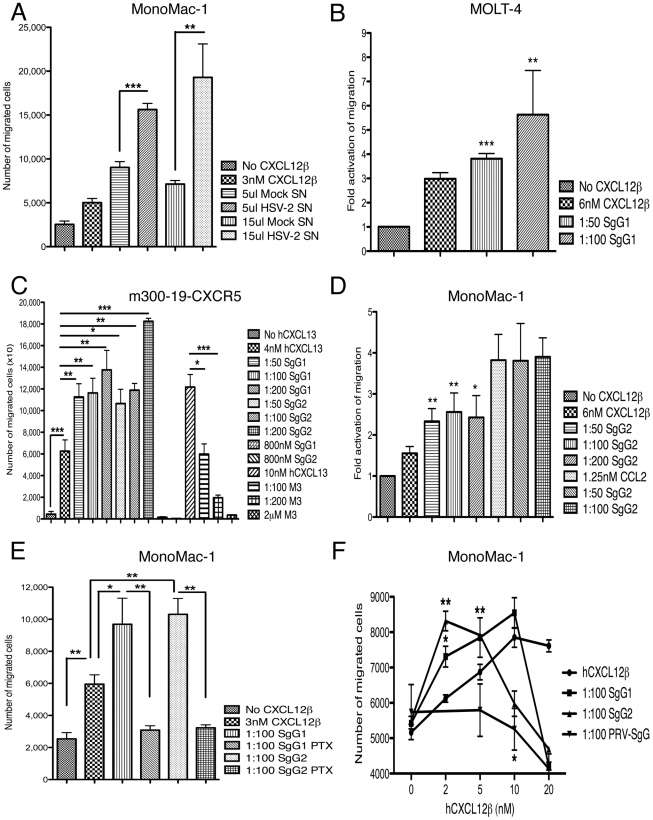
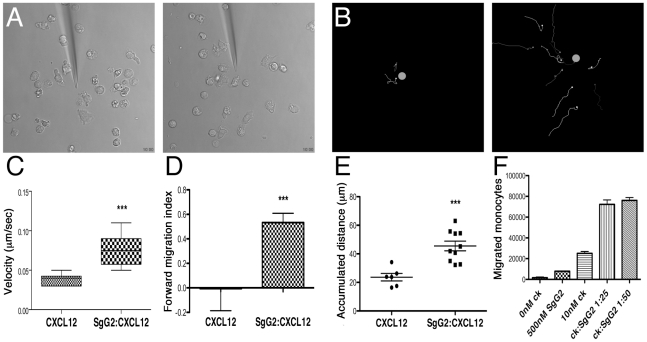
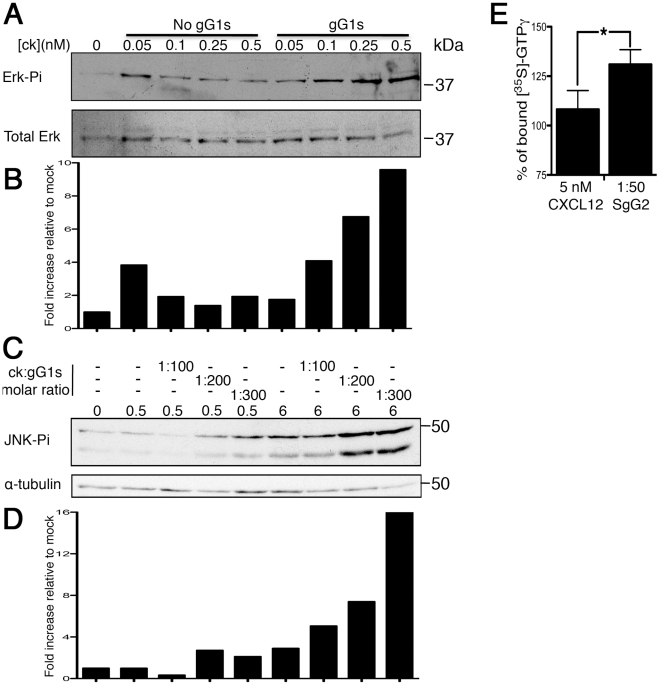
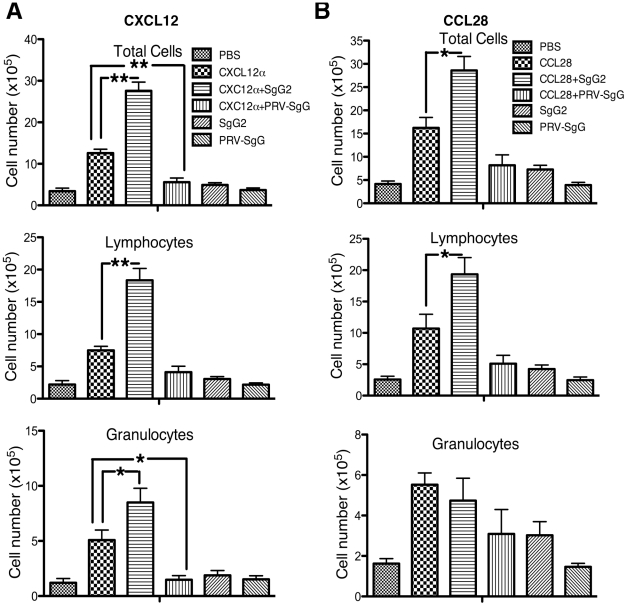
Herpes simplex virus (HSV) types 1 and 2 are highly prevalent human neurotropic pathogens that cause a variety of diseases, including lethal encephalitis. The relationship between HSV and the host immune system is one of the main determinants of the infection outcome. Chemokines play relevant roles in antiviral response and immunopathology, but the modulation of chemokine function by HSV is not well understood. We have addressed the modulation of chemokine function mediated by HSV. By using surface plasmon resonance and crosslinking assays we show that secreted glycoprotein G (SgG) from both HSV-1 and HSV-2 binds chemokines with high affinity. Chemokine binding activity was also observed in the supernatant of HSV-2 infected cells and in the plasma membrane of cells infected with HSV-1 wild type but not with a gG deficient HSV-1 mutant. Cell-binding and competition experiments indicate that the interaction takes place through the glycosaminoglycan-binding domain of the chemokine. The functional relevance of the interaction was determined both in vitro, by performing transwell assays, time-lapse microscopy, and signal transduction experiments; and in vivo, using the air pouch model of inflammation. Interestingly, and in contrast to what has been observed for previously described viral chemokine binding proteins, HSV SgGs do not inhibit chemokine function. On the contrary, HSV SgGs enhance chemotaxis both in vitro and in vivo through increasing directionality, potency and receptor signaling. This is the first report, to our knowledge, of a viral chemokine binding protein from a human pathogen that increases chemokine function and points towards a previously undescribed strategy of immune modulation mediated by viruses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Herpes simplex virus particles interact with chemokines and enhance cell migration.J Gen Virol. 2016 Nov;97(11):3007-3016. doi: 10.1099/jgv.0.000616. Epub 2016 Sep 27. J Gen Virol. 2016. PMID: 27692045
-
Secreted herpes simplex virus-2 glycoprotein G modifies NGF-TrkA signaling to attract free nerve endings to the site of infection.PLoS Pathog. 2015 Jan 22;11(1):e1004571. doi: 10.1371/journal.ppat.1004571. eCollection 2015 Jan. PLoS Pathog. 2015. PMID: 25611061 Free PMC article.
-
Herpes simplex virus enhances chemokine function through modulation of receptor trafficking and oligomerization.Nat Commun. 2015 Jan 27;6:6163. doi: 10.1038/ncomms7163. Nat Commun. 2015. PMID: 25625471
-
Animal models of herpes simplex virus immunity and pathogenesis.J Neurovirol. 2015 Feb;21(1):8-23. doi: 10.1007/s13365-014-0302-2. Epub 2014 Nov 12. J Neurovirol. 2015. PMID: 25388226 Review.
-
Herpes Simplex Virus Evasion of Early Host Antiviral Responses.Front Cell Infect Microbiol. 2019 Apr 30;9:127. doi: 10.3389/fcimb.2019.00127. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31114761 Free PMC article. Review.
Cited by
-
Herpes Simplex Virus 2 Counteracts Neurite Outgrowth Repulsion during Infection in a Nerve Growth Factor-Dependent Manner.J Virol. 2020 Sep 29;94(20):e01370-20. doi: 10.1128/JVI.01370-20. Print 2020 Sep 29. J Virol. 2020. PMID: 32669337 Free PMC article.
-
Viral modulation of type II interferon increases T cell adhesion and virus spread.Nat Commun. 2024 Jun 22;15(1):5318. doi: 10.1038/s41467-024-49657-4. Nat Commun. 2024. PMID: 38909022 Free PMC article.
-
Genome Sequence of Herpes Simplex Virus 1 Strain SC16.Genome Announc. 2017 Jan 26;5(4):e01392-16. doi: 10.1128/genomeA.01392-16. Genome Announc. 2017. PMID: 28126930 Free PMC article.
-
Targeting Chemokine-Glycosaminoglycan Interactions to Inhibit Inflammation.Front Immunol. 2020 Mar 31;11:483. doi: 10.3389/fimmu.2020.00483. eCollection 2020. Front Immunol. 2020. PMID: 32296423 Free PMC article. Review.
-
Chemokines encoded by herpesviruses.J Leukoc Biol. 2017 Nov;102(5):1199-1217. doi: 10.1189/jlb.4RU0417-145RR. Epub 2017 Aug 28. J Leukoc Biol. 2017. PMID: 28848041 Free PMC article. Review.
References
-
- Wutzler P, Doerr HW, Farber I, Eichhorn U, Helbig B, et al. Seroprevalence of herpes simplex virus type 1 and type 2 in selected German populations-relevance for the incidence of genital herpes. J Med Virol. 2000;61:201–207. - PubMed
-
- Kimberlin DW. Management of HSV encephalitis in adults and neonates: diagnosis, prognosis and treatment. Herpes. 2007;14:11–16. - PubMed
-
- Kimberlin D. Herpes simplex virus, meningitis and encephalitis in neonates. Herpes. 2004;11(Suppl 2):65A–76A. - PubMed
-
- Baggiolini M. Chemokines and leukocyte traffic. Nature. 1998;392:565–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
