

PRMT1 methylates the single Argonaute of Toxoplasma gondii and is important for the recruitment of Tudor nuclease for target RNA cleavage by antisense guide RNA
- PMID: 22309152
- PMCID: PMC3682492
- DOI: 10.1111/j.1462-5822.2012.01763.x
PRMT1 methylates the single Argonaute of Toxoplasma gondii and is important for the recruitment of Tudor nuclease for target RNA cleavage by antisense guide RNA
Abstract
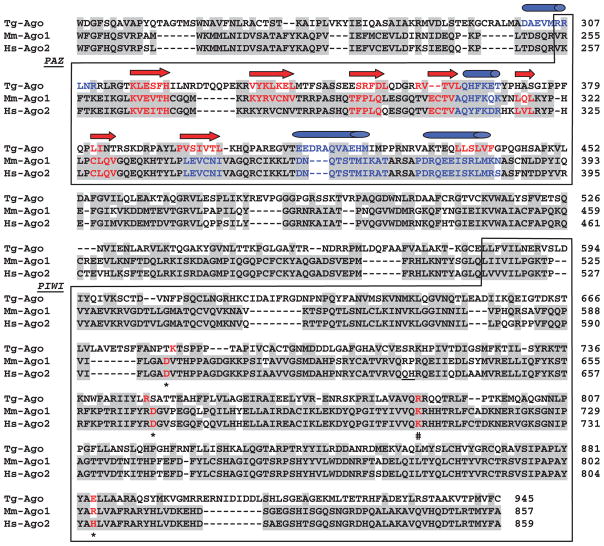
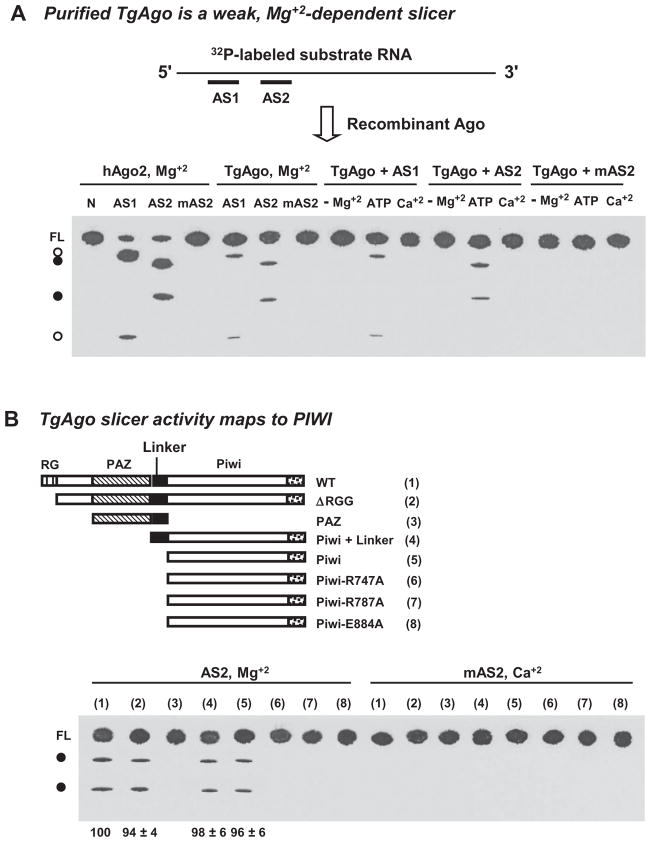
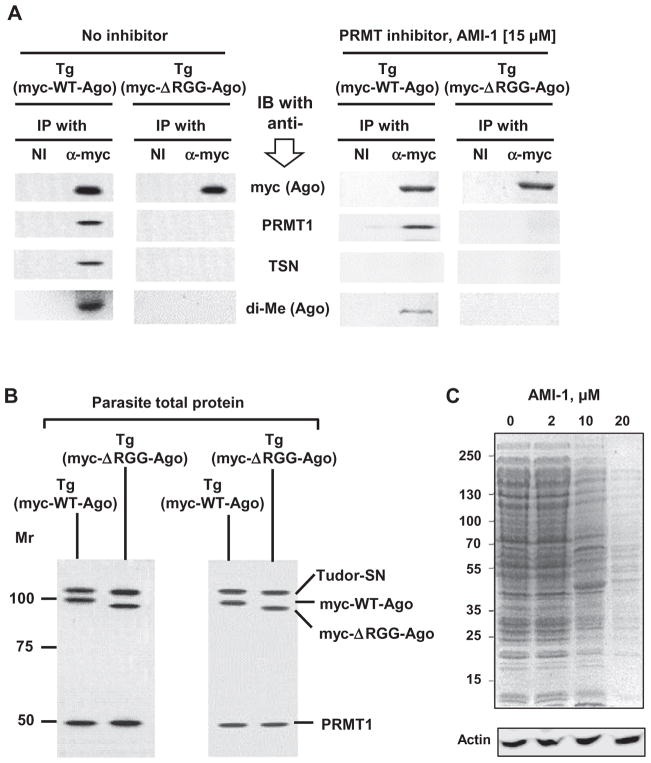
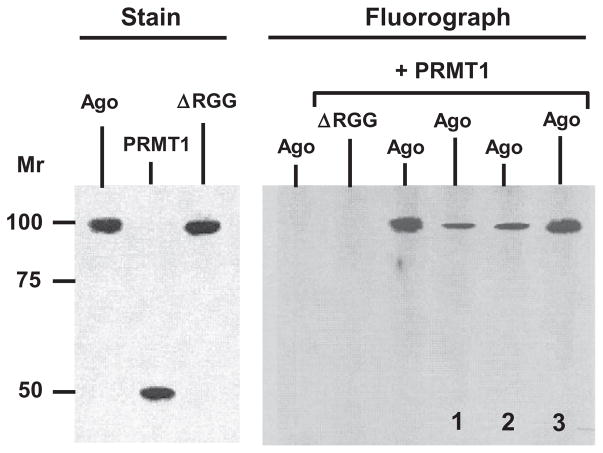
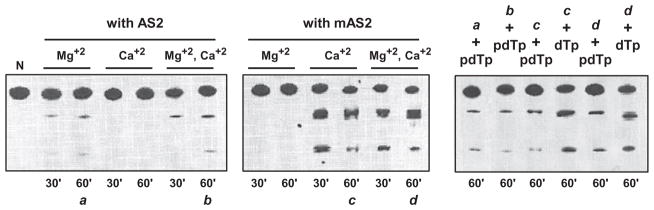
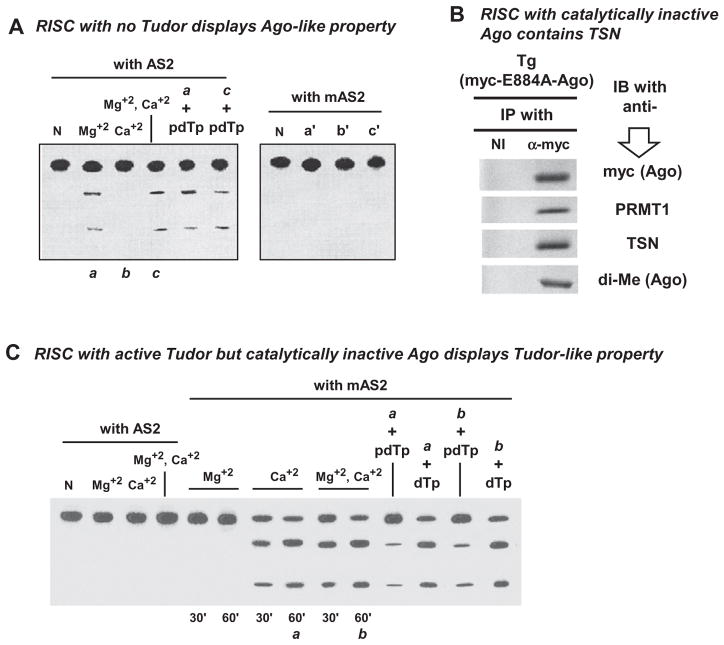
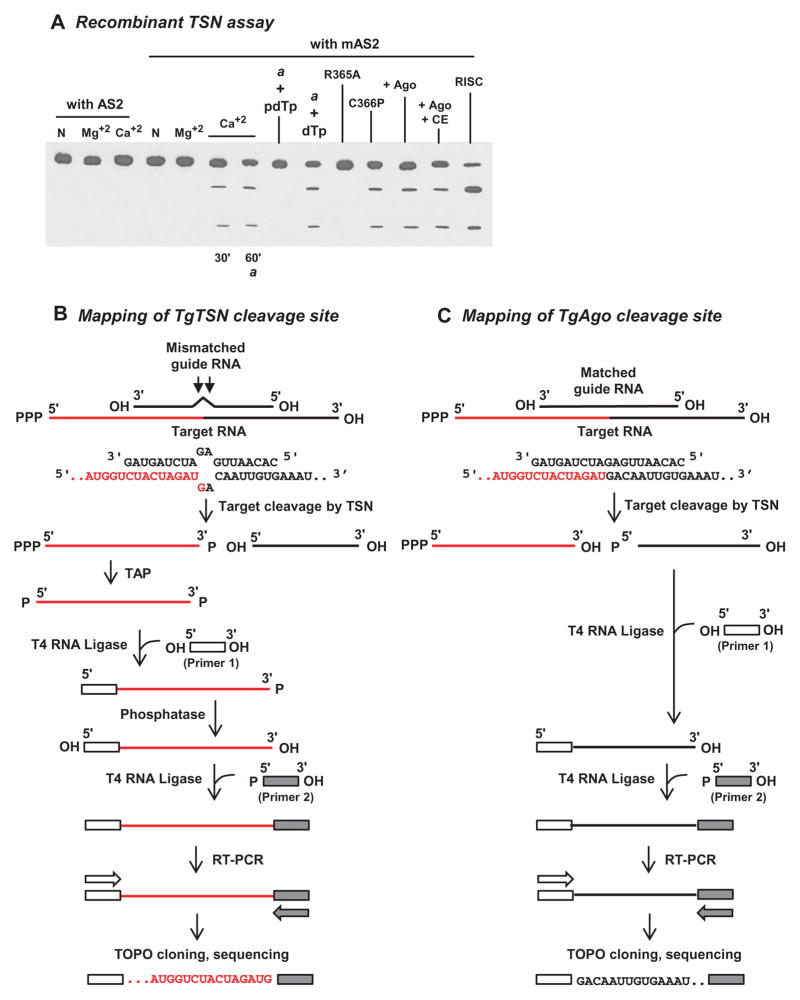
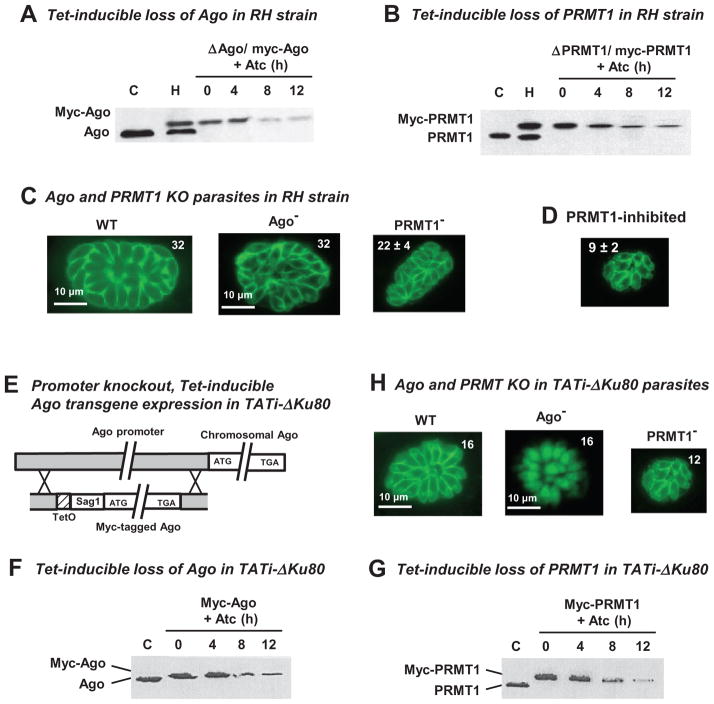
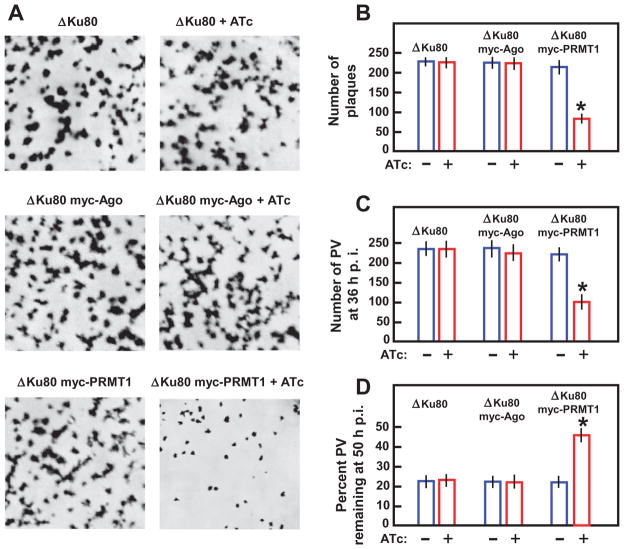
Argonaute (Ago) plays a central role in RNA interference in metazoans, but its status in lower organisms remains ill-defined. We report on the Ago complex of the unicellular protozoan, Toxoplasma gondii (Tg), an obligatory pathogen of mammalian hosts. The PIWI-like domain of TgAgo lacked the canonical DDE/H catalytic triad, explaining its weak target RNA cleavage activity. However, TgAgo associated with a stronger RNA slicer, a Tudor staphylococcal nuclease (TSN), and with a protein Arg methyl transferase, PRMT1. Mutational analysis suggested that the N-terminal RGG-repeat domain of TgAgo was methylated by PRMT1, correlating with the recruitment of TSN. The slicer activity of TgAgo was Mg(2+)-dependent and required perfect complementarity between the guide RNA and the target. In contrast, the TSN activity was Ca(2+) -dependent and required an imperfectly paired guide RNA. Ago knockout parasites showed essentially normal growth, but in contrast, the PRMT1 knockouts grew abnormally. Chemical inhibition of Arg-methylation also had an anti-parasitic effect. These results suggest that the parasitic PRMT1 plays multiple roles, and its loss affects the recruitment of a more potent second slicer to the parasitic RNA silencing complex, the exact mechanism of which remains to be determined.
© 2012 Blackwell Publishing Ltd.
Figures

Similar articles
-
Single argonaute protein from Toxoplasma gondii is involved in the double-stranded RNA induced gene silencing.Int J Parasitol. 2006 Aug;36(9):1003-14. doi: 10.1016/j.ijpara.2006.04.014. Epub 2006 May 30. Int J Parasitol. 2006. PMID: 16753169
-
Comparative Monomethylarginine Proteomics Suggests that Protein Arginine Methyltransferase 1 (PRMT1) is a Significant Contributor to Arginine Monomethylation in Toxoplasma gondii.Mol Cell Proteomics. 2017 Apr;16(4):567-580. doi: 10.1074/mcp.M117.066951. Epub 2017 Jan 31. Mol Cell Proteomics. 2017. PMID: 28143887 Free PMC article.
-
Crystal structure of A. aeolicus argonaute, a site-specific DNA-guided endoribonuclease, provides insights into RISC-mediated mRNA cleavage.Mol Cell. 2005 Aug 5;19(3):405-19. doi: 10.1016/j.molcel.2005.07.011. Mol Cell. 2005. PMID: 16061186 Free PMC article.
-
Protein arginine methylation in parasitic protozoa.Eukaryot Cell. 2011 Aug;10(8):1013-22. doi: 10.1128/EC.05103-11. Epub 2011 Jun 17. Eukaryot Cell. 2011. PMID: 21685318 Free PMC article. Review.
-
When Argonaute takes out the ribonuclease sword.J Biol Chem. 2024 Jan;300(1):105499. doi: 10.1016/j.jbc.2023.105499. Epub 2023 Nov 27. J Biol Chem. 2024. PMID: 38029964 Free PMC article. Review.
Cited by
-
Protein arginine methyltransferases in protozoan parasites.Parasitology. 2022 Apr;149(4):427-435. doi: 10.1017/S0031182021002043. Epub 2021 Dec 6. Parasitology. 2022. PMID: 35331350 Free PMC article. Review.
-
The unique dual targeting of AGO1 by two types of PRMT enzymes promotes phasiRNA loading in Arabidopsis thaliana.Nucleic Acids Res. 2024 Mar 21;52(5):2480-2497. doi: 10.1093/nar/gkae045. Nucleic Acids Res. 2024. PMID: 38321923 Free PMC article.
-
R-Methylation in Plants: A Key Regulator of Plant Development and Response to the Environment.Int J Mol Sci. 2024 Sep 14;25(18):9937. doi: 10.3390/ijms25189937. Int J Mol Sci. 2024. PMID: 39337424 Free PMC article. Review.
-
Induction of interferon-stimulated genes by IRF3 promotes replication of Toxoplasma gondii.PLoS Pathog. 2015 Mar 26;11(3):e1004779. doi: 10.1371/journal.ppat.1004779. eCollection 2015 Mar. PLoS Pathog. 2015. Retraction in: PLoS Pathog. 2024 May 31;20(5):e1012264. doi: 10.1371/journal.ppat.1012264 PMID: 25811886 Free PMC article. Retracted.
-
Entamoeba histolytica: protein arginine transferase 1a methylates arginine residues and potentially modify the H4 histone.Parasit Vectors. 2015 Apr 10;8:219. doi: 10.1186/s13071-015-0820-7. Parasit Vectors. 2015. PMID: 25889855 Free PMC article.
References
-
- Al Riyahi A, Al-Anouti F, Al-Rayes M, Ananvoranich S. Single argonaute protein from Toxoplasma gondii is involved in the double-stranded RNA induced gene silencing. Int J Parasitol. 2006;36:1003–1014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
