

Review
doi: 10.1016/j.coi.2011.12.006.
Epub 2012 Feb 3.
A new eye on NLR proteins: focused on clarity or diffused by complexity?
Affiliations
- PMID: 22305607
- PMCID: PMC3482489
- DOI: 10.1016/j.coi.2011.12.006
Item in Clipboard
Review
A new eye on NLR proteins: focused on clarity or diffused by complexity?
Curr Opin Immunol.
2012 Feb.
Abstract
The nucleotide-binding domain leucine-rich repeat proteins (NLRs) represent the major class of intracellular innate immune receptors in plants and animals. Understanding their functions is a major challenge in immunology. This review highlights recent efforts toward elucidating NLR functions in human and plants. We compare unconventional aspects of NLR proteins across the two kingdoms. We review recent advances describing P-loop independent activation, nuclear-cytoplasmic trafficking, oligomerization and multimerization requirements for signaling, and for expanded functions beyond pathogen recognition by several NLR proteins.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures

Architecture of NLR proteins. Domain organization of (a) plant NLRs and (b) animal NLRs is depicted and representative members of each category are shown. (c) The conserved domains of NLR proteins are represented. N-terminal coiled-coil (CC) and Toll/interleukin-1 (TIR) and C-terminal WRKY domains are specific to plants, whereas N-terminal baculoviral inhibitory repeat (BIR) domain, caspase recruitment domain (CARD), pyrin domain (PYD), activation domain (AD), and undefined domains are specific to animals.
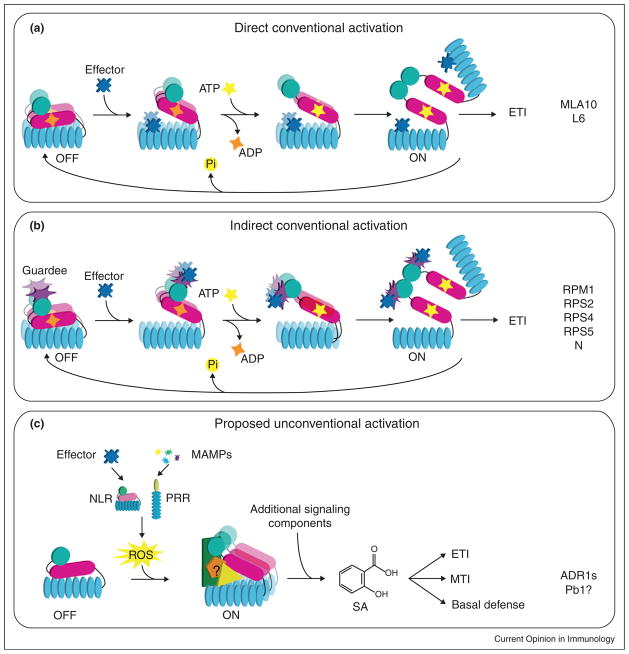
Model for activation of plant NLR proteins. (a and b) NLRs are normally in an ‘off’ state that is achieved by the inhibitory function of the LRR domain preventing the protein activation via the NB molecular switch. The NLR can either be a dimer (MLA10) or a monomer (L6) before activation. NLR proteins can recognize the effector protein directly through the LRR domain (a), or through modifications of the host target of an effector that typically interacts with the NLR N-terminal domain (b). Recognition of the pathogen effector triggers the release of the inhibitory LRR domain and this conformational change allows for exchange of ADP to ATP. Binding of ATP results in a second conformational change that allows the N-terminal domains to physically interact, thus activating the NLR. Based on the dimerization of NOD1 through the CARD domains, this mechanism is likely to be conserved in animal NLRs. (c) When a P-loop is not required for NLR function, we propose that the NLR is activated by an oxidative burst resulting from either NLR-mediated or PRR-mediated effector-dependent or MAMP-dependent recognition. Unconventional NLRs could function as scaffolds for interactions with unknown partners. Heterotypic or homotypic interactions with accessory proteins or other NLRs could result in the activation of downstream signaling which leads to ETI, MTI, and basal defense.
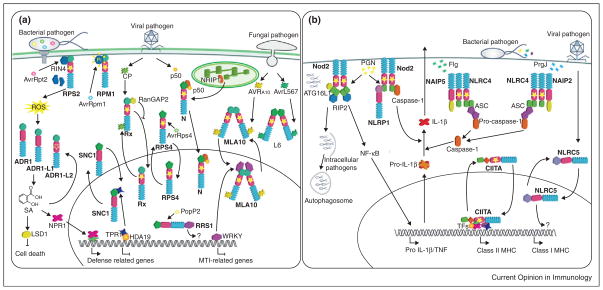
NLR functions in plant and animal cells. (a) In plants, exposure to pathogen effectors results in activation of NLR proteins. NLRs can either be localized at the plasma membrane, the cytosol, or shuttle between the cytosol and the nucleus. Recognition of effectors can either be direct (L6, MLA10, Rx) or mediated by host proteins (RPM1, RPS2, N). RPM1 and RPS2 recognize the bacterial effectors AvrRpm1 and AvrRpt2 respectively through either phosphorylation or cleavage of the host protein RIN4. N recognizes the 50 kDa helicase (p50) domain of Tobacco Mosaic Virus, which recruits the chloroplastic protein NRIP1 to form an immune receptor complex through N TIR domain. Activation of MLA10 and L6 following recognition of AVRA10 and AvrL567through their respective LRR domains results in homodimerization through their N-terminal domains. Rx is activated by recognition of the Potato Virus X coat protein (CP) and activation results in the inhibition of the cytoplasmic retention protein RanGAP2. SNC1 shuttles from the cytoplasm to the nucleus upon activation from a yet unknown signal. In the nucleus SNC1 recruits the transcriptional co-repressor TPR1, which normally associates to histone deacetylase 19 (HDA19) to repress transcription of defense-related genes. RRS1 is a TIR-NLR that carries a WRKY domain at its C-terminus and localizes to the nucleus. PopP2 is an effector from Ralstonia solanacearum that associates to RRS1 in the nucleus. Transcriptional activity for the WRKY domain of RRS1 has not yet been reported. ADR1 NLRs are activated downstream of an oxidative burst derived from either effector-mediated activation of unrelated NLRs or MAMP recognition. ADR1 proteins are regulators of SA accumulation and SA levels are controlled by LSD1, a negative regulator of cell death. SA is essential for Non-expressor or PR genes 1 (NPR1) function to transcriptionally regulate defense-related genes. (b) In animals MAMPs are perceived by cytoplasmic NLRs, however Nod2 is functional at the plasma membrane where it perceives PGN. Nod2 responds to PGN and its activation triggers the recruitment of the adaptor protein RIP2, resulting in downstream signaling and the expression of pro-inflammatory cytokines. Moreover, Nod2 recruits the autophagy protein ATG16L to the plasma membrane at the bacterial entry site in a mechanism that does not require RIP2. Additionally, Nod2 acts in synergy with NLRP1 in PNG sensing and subsequent caspase-1 activation, thus cleavage of IL-1β zymogen. The NLRC4 inflammasome is activated in response to the bacterial proteins flagellin (flg) and PrgJ. The bacterial ligands trigger oligomerization of NLRC4 with the bacterial sensor NAIP NLRs. The NAIP2-NLRC4 complex confers specificity for PrgJ recognition, whereas NAIP5-NLRC4 to flg. In both cases activation of the downstream signaling results in caspase-1 activation and cytokines secretion via recruitment of the adaptor protein ASC (Apoptosis-associated Speck-like Protein Containing a CARD). CIITA and NLRC5 shuttle between the cytoplasm and the nucleus and function as transcriptional regulators of genes encoding the major histocompatibility complex I and II. CIITA requires its GTPase activity to access the nucleus, thus to recruit transcription factors (TFs) and histone modifying enzymes. Filled stars indicate a requirement of an intact P-loop for the NLR function, an empty star indicates that an intact P-loop is dispensable for the function, as suggested by mutagenesis and/or biochemical analysis.
Similar articles
-
NLR, the nucleotide-binding domain leucine-rich repeat containing gene family.Curr Opin Immunol. 2008 Feb;20(1):3-9. doi: 10.1016/j.coi.2008.01.003. Epub 2008 Feb 15. Curr Opin Immunol. 2008. PMID: 18280719 Review.
-
NOD-like receptor cooperativity in effector-triggered immunity.Trends Immunol. 2014 Nov;35(11):562-70. doi: 10.1016/j.it.2014.09.005. Epub 2014 Oct 9. Trends Immunol. 2014. PMID: 25308923 Review.
-
Signal transduction pathways used by NLR-type innate immune receptors.Mol Biosyst. 2008 May;4(5):380-6. doi: 10.1039/b718948f. Epub 2008 Mar 27. Mol Biosyst. 2008. PMID: 18414735 Review.
-
The microbial and danger signals that activate Nod-like receptors.Cytokine. 2008 Sep;43(3):368-73. doi: 10.1016/j.cyto.2008.07.013. Epub 2008 Aug 19. Cytokine. 2008. PMID: 18715799 Review.
-
The HSP90-SGT1 chaperone complex for NLR immune sensors.Annu Rev Plant Biol. 2009;60:139-64. doi: 10.1146/annurev.arplant.59.032607.092906. Annu Rev Plant Biol. 2009. PMID: 19014346 Review.
Cited by
-
Multiple Evolutionary Events Involved in Maintaining Homologs of Resistance to Powdery Mildew 8 in Brassica napus.Front Plant Sci. 2016 Jul 21;7:1065. doi: 10.3389/fpls.2016.01065. eCollection 2016. Front Plant Sci. 2016. PMID: 27493652 Free PMC article.
-
Advances in Biological Control and Resistance Genes of Brassicaceae Clubroot Disease-The Study Case of China.Int J Mol Sci. 2023 Jan 2;24(1):785. doi: 10.3390/ijms24010785. Int J Mol Sci. 2023. PMID: 36614228 Free PMC article. Review.
-
Genome-Wide Identification, Characterization, and Comparative Analysis of NLR Resistance Genes in Coffea spp.Front Plant Sci. 2022 Jul 7;13:868581. doi: 10.3389/fpls.2022.868581. eCollection 2022. Front Plant Sci. 2022. PMID: 35874027 Free PMC article.
-
Identification of mildew resistance in wild and cultivated Central Asian grape germplasm.BMC Plant Biol. 2013 Oct 4;13:149. doi: 10.1186/1471-2229-13-149. BMC Plant Biol. 2013. PMID: 24093598 Free PMC article.
-
The nuclear immune receptor RPS4 is required for RRS1SLH1-dependent constitutive defense activation in Arabidopsis thaliana.PLoS Genet. 2014 Oct 23;10(10):e1004655. doi: 10.1371/journal.pgen.1004655. eCollection 2014 Oct. PLoS Genet. 2014. PMID: 25340333 Free PMC article.
References
-
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
-
- Segonzac C, Zipfel C. Activation of plant pattern-recognition receptors by bacteria. Curr Opin Microbiol. 2011;14:54–61. - PubMed
-
- Guo YL, Fitz J, Schneeberger K, Ossowski S, Cao J, Weigel D. Genome-wide comparison of NB-LRR encoding genes in Arabidopsis. Plant Physiol. 2011;157:757–769. The authors compared the repertoire of NLR proteins of Arabidopsis thaliana with its outcrossing relative A. lyrata and analyzed inter-specific and intra-specific diversity. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
