

doi: 10.1199/tab.0123.
Epub 2009 Sep 11.
Peroxisome biogenesis and function
- PMID: 22303249
- PMCID: PMC3243405
- DOI: 10.1199/tab.0123
Item in Clipboard
Peroxisome biogenesis and function
Arabidopsis Book.
2009.
Abstract
Peroxisomes are small and single membrane-delimited organelles that execute numerous metabolic reactions and have pivotal roles in plant growth and development. In recent years, forward and reverse genetic studies along with biochemical and cell biological analyses in Arabidopsis have enabled researchers to identify many peroxisome proteins and elucidate their functions. This review focuses on the advances in our understanding of peroxisome biogenesis and metabolism, and further explores the contribution of large-scale analysis, such as in sillco predictions and proteomics, in augmenting our knowledge of peroxisome function In Arabidopsis.
Figures
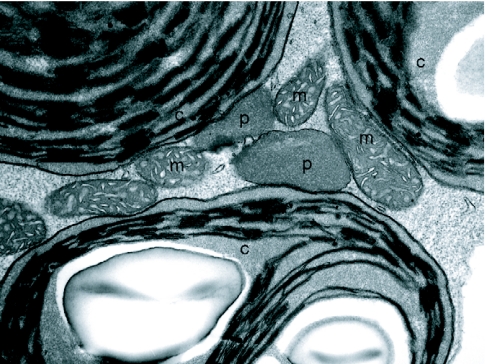
Transmission electron micrograph of organelles from a green cotyledon cell in a 7d wild-type Arabidopsis seedling. The association of peroxisomes (p) with chloroplasts (c) and mitochondria (m) are shown.
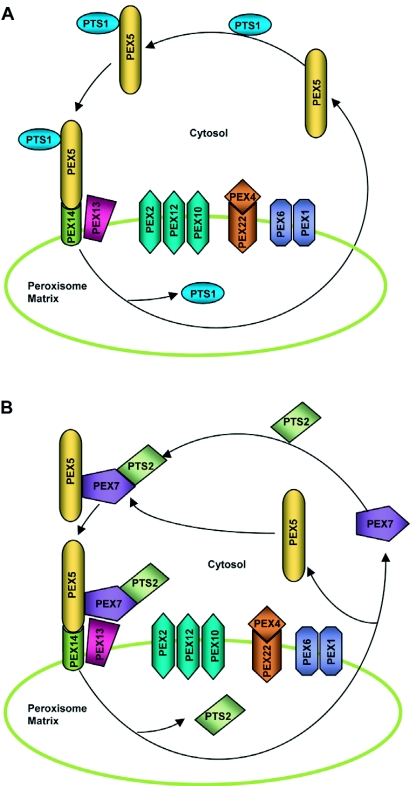
Speculative model for matrix protein import. (A) PTS1 import. PEX5 recognizes and binds PTS1-containing proteins in the cytosol. The receptor-PTS1 protein complex then traffics to the peroxisome where it associates with the docking complex on the peroxisome membrane. The docking complex in Arabidopsis probably comprises PEX14 and PEX13 and is believed to tether the receptor-protein complex to the peroxisome membrane through the interaction of PEX5 with PEX14. Subsequently, the PTS1 protein is dissociated from PEX5 and released into the peroxisomal matrix by an unknown mechanism. The export of the receptor to the cytosol might be facilitated by its possible ubiquitination (not shown) by the PEX22-anchored PEX4, a putative ubiquitin-conjugating enzyme, followed by ATP-driven dislocation mediated by the peroxisomal AAA ATPases PEX1 and PEX6. The RING complex, which is composed of the PEX2, PEX10 and PEX12 RING peroxins, plausibly plays a role in the import and export processes, although the exact function(s) of this complex are not well understood. (B) PTS2 import. PEX7 recognizes and binds PTS2-containing proteins in the cytosol. The receptor-PTS2 protein complex binds coordinately with PEX5 and is ferried to the peroxisome docking complex. The subsequent steps of import are assumed to be similar to PTS1 import. The events facilitating the release of PEX7 and PTS2 protein are not well known.
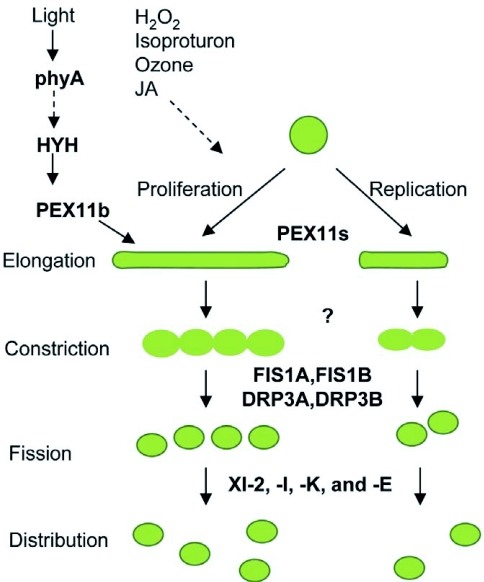
Model for peroxisome proliferation and replication in Arabidopsis. The PEX11 family of proteins (PEX11a-e) acts in the initial steps of peroxisome proliferation and replication, resulting in pronounced elongation or expansion of peroxisomes. Proteins overseeing the subsequent membrane constriction are not known. Fission of the constricted peroxisomes is enabled by the scission activities of dynamin-related proteins DRP3A and DRP3B, which are tethered to the peroxisome membrane by FIS1A and FIS1B. The divided peroxisomes are then transported to various parts of the cell by the indicated myosin proteins (XI-2, XI-I, XI-E, XI-K) along actin cables. The transcription factor HYH has been implicated in the phyA-mediated light regulation of peroxisome elongation via activation of the PEX11b gene. Hydrogen peroxide (H2O2), isoproturon, ozone and Jasmonates (JA) are other cues that induce or repress peroxisome proliferation through factors currently unknown, indicated by dotted arrow.
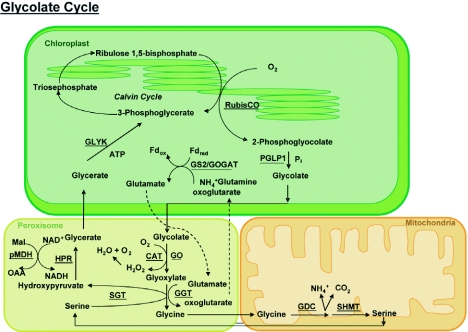
Schematic of the photorespiratory pathway (for details see text in section 3.4).
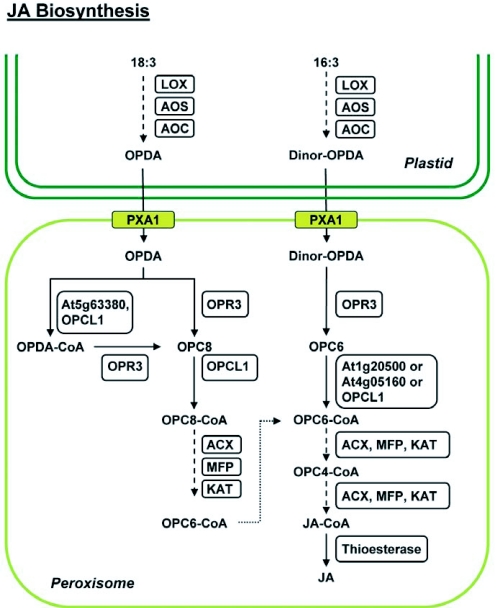
JA biosynthesis pathway. Biosynthesis of (+)-7-iso-jasmonic acid (JA) initiates in the chloroplast with the sequential action of lipoxygenase (LOX), allene oxide synthase (AOS), and allene oxide cyclase (AOC) on linolenic acid (C18:3) or hexadeca-trienoic acid (C16:3) to generate 12-oxophytodienoic acid (OPDA) and dinor-OPDA, respectively. Transport of OPDA and dinor-OPDA into peroxisomes is facilitated either by the ABC transporter (PXA1) or by an uncharacterized pathway (not shown). Oxophytodienoic acid reductase 3 (OPR3) reduces OPDA to 3-oxo-2-(2′-[Z]-pentenyl) cyclopentane-1-octanoic acid (OPC8) and dnOPDA to 3-oxo-2-(2′-pentenyl)-cyclopentane-1-hexanoic acid (OPC6), with the resultant compounds being activated to their corresponding CoA esters by OPC:8 CoA ligasel (OPCL1) or the indicated acyl-CoA synthetases (At1g20500, At4g05160, At5g63380). The CoA derivatives undergo β-oxidation by the consecutive activities of acyl-CoA oxidase (ACX), multifunctional protein (MFP) and 3-ketoacyl-CoA thiolase (KAT), eventually generating JA-CoA after being subjected to the requisite number of β-oxidation cycles (3 for OPCS-CoA and 2 for OPC6-CoA). A putative thioesterase cleaves the CoA moiety, releasing JA. Dotted arrows depict catalysis of substrate(s) by the consecutive actions of the indicated enzymes.
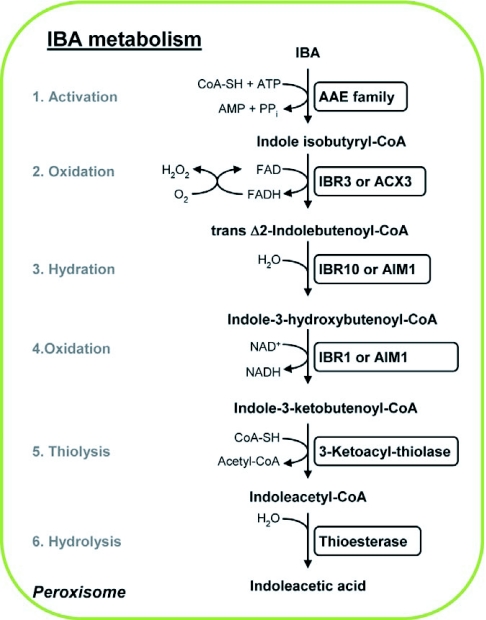
IBA metabolism pathway in the peroxisome. An uncharacterized acyl-activating enzyme (AAE) activates IBA by the addition of a CoA moiety. The IBA-CoA derivative goes through a single β-oxidation cycle, resulting in its sequential oxidation, hydration, oxidation, and thiolysis to yield IAA-CoA and an acetyl-CoA. The putative enzymes involved in each of these catalytic steps are indicated. The IAA-CoA thus generated is hydrolysed by the activity of an unknown thioesterase to release the active auxin IAA. The conversion of 2,4-DB to 2,4-D most likely follows the same cascade of enzymatic reactions differing only in the initial step, where AAE18 probably catalyzes the activation of 2,4-DB. For further details see text in section 3.6.
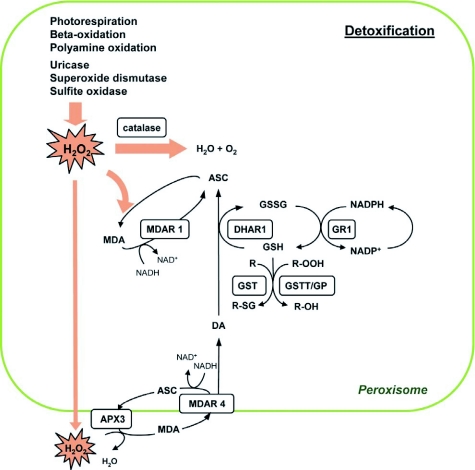
Model for detoxification reactions in plant peroxisomes. H2O2 is a potent ROS generated as a by-product of various metabolic reactions occurring within the peroxisome. It has the potential to inflict severe damage to enzymes and membrane lipids, thus necessitating its rapid detoxification. H2O2 is scavenged either by the catalases (CAT1, -2, and -3) in the matrix or through the activity of the ascorbate (ASC)-glutathione (GSH) cycle. In the latter case, H2O2 is reduced to H2O by ascorbate peroxidase (APX), producing monodehydroascorbate (MDA). MDA can either be reduced to ASC by monodehydroascorbate reductase (MDAR) with the simultaneous oxidation of NADH to NAD+ or, alternatively, MDA undergoes disproportionation to yield ASC and dehydroascorbate (DA). MDAR isoforms are associated both with the peroxisomal matrix (MDAR1) and the peroxisome membrane (MDAR4). Dehydroascorbate reductase (DHAR) acts on DA, utilizing reduced glutathione (GSH) to reduce DA to ASC; GSH is oxidized to GSSG in this reaction. Glutathione reductase (GR) regenerates GSH by using the reduced cofactor NADPH. Peroxisomal isocitrate dehydrogenase and 6-phosphogluconate dehydrogenase probably catalyze the reduction of NADP+ to NADPH, regenerating the reduced cofactor for the operation of GR (not shown). Three members of the theta family of glutathione S-transferases (GSTT) function mainly as glutathione peroxidases (GP), reducing fatty acid hydroperoxides (R-OOH) and H2O2 at the expense of GSH, although they also show limited transferase activity in conjugating GSH to metabolite (R).
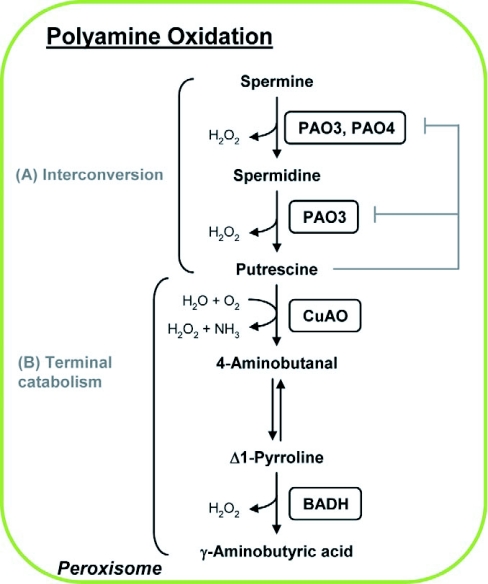
Polyamine oxidation pathways in peroxisomes. (A) Interconversion pathway. Spermine and spermidine are substrates for peroxisomal polyamine oxidases (PAO2-4). PAO4 acts specifically on spermine to oxidize it to spermidine. PAO3 catalyses the sequential back-conversion of spermine to spermidine and spermidine to putrescine, and is subjected to feedback inhibition by its end product, putrescine. The specific substrate for PAO2 has not been established. (B) Speculative pathway for terminal catabolism. Peroxisomal copper amine oxidase (CuAO) oxidatively deaminates putrescine to produce ammonia and H2O2. The reaction product 4-aminobutyraldehyde spontaneously isomerizes to Δ1-pyrroline, which is a substrate for peroxisomal betaine aldehyde dehydrogenase (BADH) that catalyzes its conversion to γ-aminobutyric acid (GABA) and releases H2O2 as a by-product.
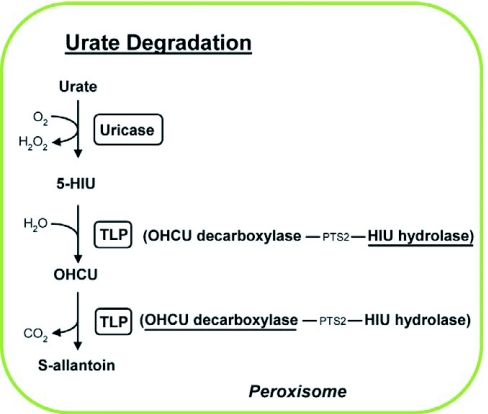
Urate degradation The oxidation of urate is catalyzed by uricase to generate 5-Hydroxyisourate (5-HIU). 5-HIU is subsequently hydrolysed by HIU hydrolase (HIUase) to yield 2-oxo-4-hydroxy-4-carboxy-5-ureidoimidazoline (OHCU), which undergoes decarboxylation by OHCU decarboxylase to release carbon dioxide and produce S-allantoin. The latter two activities are imparted by the bifunctional transthyretin-like protein (TLP) in Arabidopsis.
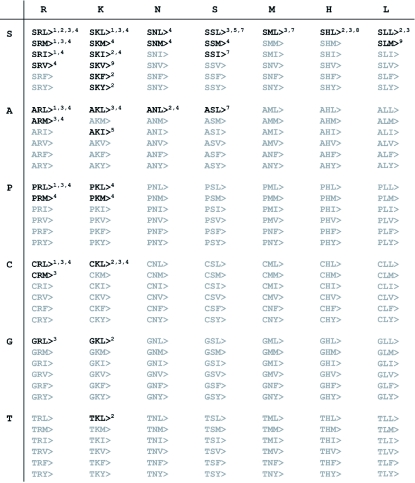
Summary of known plant PTS1 tripeptides. The PTS1 tripeptides that have been experimentally verified as functional PTS or found in significant numbers (≥2) in datasets of homologous sequences (Reumann, 2004) are in bold. All other amino acid combinations of amino acid residues present in PTS1 tripeptides are in grey. SQL> and SAL>, both of which have been characterized only in PEX5 protein-peptide interaction studies (Kragler et al., 1998) and carry amino acid residues (Q and A at pos. -2) that have not been described in plant in vivo studies are not shown. References 1–9 are: Hayashi et al., 1997, Mullen et al., 1997, Kragler et al., 1998, Reumann, 2004, Lisenbee et al., 2005, Goepfert et al., 2006, Reumann et al., 2007, Ma and Reumann, 2008, and Reumann et al., 2009.
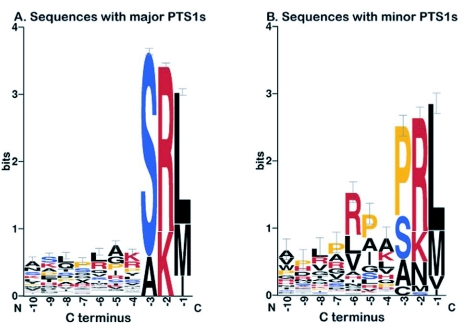
Sequence logos for plant proteins with major or minor PTS1 tripeptides. Position-specific amino acid compositions of the C-terminal 10 amino acid residues of true peroxisomal PTS1-containing proteins, which carry either one of the nine major PTS1s (A) or one of the eleven minor PTS1s (B) as defined by Reumann (2004), are shown. The sequence logo was generated by WebLogo 3 (weblogo.berkeley.edu/ ).
Similar articles
-
Dynamics of Peroxisome Homeostasis and Its Role in Stress Response and Signaling in Plants.Front Plant Sci. 2019 Jun 4;10:705. doi: 10.3389/fpls.2019.00705. eCollection 2019. Front Plant Sci. 2019. PMID: 31214223 Free PMC article. Review.
-
Proteome of Plant Peroxisomes.Subcell Biochem. 2018;89:3-45. doi: 10.1007/978-981-13-2233-4_1. Subcell Biochem. 2018. PMID: 30378017 Review.
-
Novel proteins, putative membrane transporters, and an integrated metabolic network are revealed by quantitative proteomic analysis of Arabidopsis cell culture peroxisomes.Plant Physiol. 2008 Dec;148(4):1809-29. doi: 10.1104/pp.108.129999. Epub 2008 Oct 17. Plant Physiol. 2008. PMID: 18931141 Free PMC article.
-
Plant peroxisomes.Vitam Horm. 2005;72:111-54. doi: 10.1016/S0083-6729(05)72004-5. Vitam Horm. 2005. PMID: 16492470
-
Peroxisomes: versatile organelles with diverse roles in plants.New Phytol. 2020 Feb;225(4):1410-1427. doi: 10.1111/nph.16134. Epub 2019 Sep 26. New Phytol. 2020. PMID: 31442305 Review.
Cited by
-
Iron homeostasis in Arabidopsis thaliana: transcriptomic analyses reveal novel FIT-regulated genes, iron deficiency marker genes and functional gene networks.BMC Plant Biol. 2016 Oct 3;16(1):211. doi: 10.1186/s12870-016-0899-9. BMC Plant Biol. 2016. PMID: 27716045 Free PMC article.
-
Branched-Chain Volatiles in Fruit: A Molecular Perspective.Front Plant Sci. 2022 Jan 27;12:814138. doi: 10.3389/fpls.2021.814138. eCollection 2021. Front Plant Sci. 2022. PMID: 35154212 Free PMC article. Review.
-
Dynamics of Peroxisome Homeostasis and Its Role in Stress Response and Signaling in Plants.Front Plant Sci. 2019 Jun 4;10:705. doi: 10.3389/fpls.2019.00705. eCollection 2019. Front Plant Sci. 2019. PMID: 31214223 Free PMC article. Review.
-
Overexpression of Peroxisome-Localized GmABCA7 Promotes Seed Germination in Arabidopsis thaliana.Int J Mol Sci. 2022 Feb 21;23(4):2389. doi: 10.3390/ijms23042389. Int J Mol Sci. 2022. PMID: 35216505 Free PMC article.
-
Sugar Starvation Disrupts Lipid Breakdown by Inducing Autophagy in Embryonic Axes of Lupin (Lupinus spp.) Germinating Seeds.Int J Mol Sci. 2023 Jul 21;24(14):11773. doi: 10.3390/ijms241411773. Int J Mol Sci. 2023. PMID: 37511532 Free PMC article.
References
-
- Adam Z., Clarke A.K. Cutting edge of chloroplast proteolysis. Trends Plant Sci. 2002;77(1):451–456. - PubMed
-
- Adham A.R., Zolman B.K., Millius A., Bartel B. Mutations in Arabidopsis acyl-CoA oxidase genes reveal distinct and overlapping roles in beta-oxidation. Plant J. 2005;417(1):859–874. - PubMed
-
- Afitlhile M.M., Fukushige H., Nishimura M., Hildebrand D.F. A defect in glyoxysomal fatty acid beta-oxidation reduces jasmonic acid accumulation in Arabidopsis. Plant Physiol. Biochem. 2005;437(1):603–609. - PubMed
-
- Alcazar R., Marco F., Cuevas J.C., Patron M., Ferrando A., Carrasco P., Tiburcio A.F., Altabella T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006;287(1):1867–1876. - PubMed
-
- Anton M., Passreiter M., Lay D., Thai T.P., Gorgas K., Just W.W. ARF- and coatomer-mediated peroxisomal vesiculation. Cell Biochem. Biophys. 2000;327(1):27–36. Spring. - PubMed
LinkOut - more resources
Full Text Sources