

doi: 10.1199/tab.0060.
Epub 2002 Apr 4.
Root hairs
- PMID: 22303213
- PMCID: PMC3243358
- DOI: 10.1199/tab.0060
Item in Clipboard
Root hairs
Arabidopsis Book.
2002.
No abstract available
Figures

Scanning electron micrograph of a root hair cell. The hair produced by this cell is approximately 1/3 of its final length.
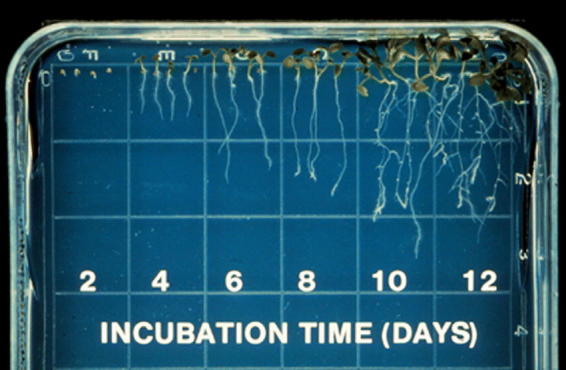
Development of Arabidopsis seedlings growing on agarose-solidified nutrient medium in vertically-oriented Petri plates. The roots grow along the surface of the medium, and root hairs are visualized easily using a low-magnification microscope.
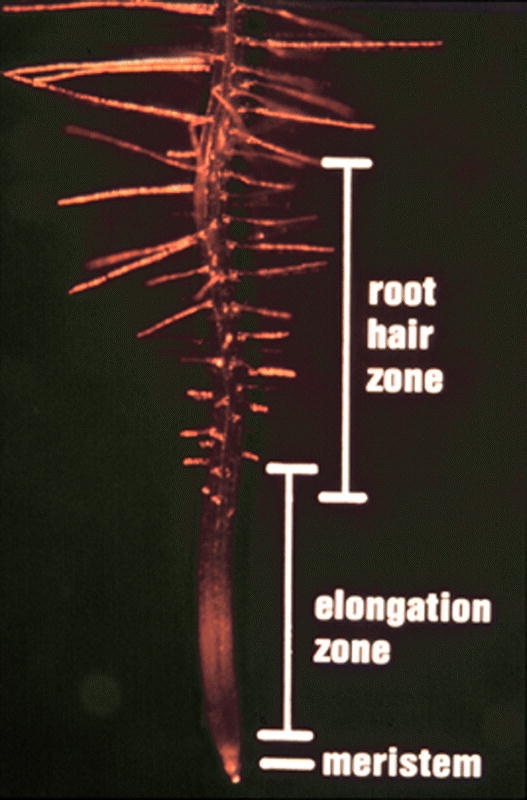
Photograph of a root tip showing the progressive development of root hair cells.
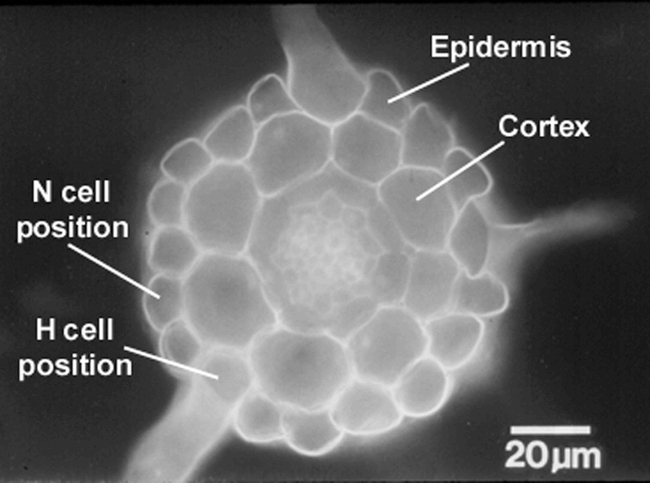
Transverse section of an Arabidopsis root, showing the position-dependent pattern of hair cells and non-hair cells. The hair-bearing cells are located outside the space separating two cortical cells (the H cell position), whereas the non-hair cells are located outside a single cortical cell (the N cell position). Three hairs are visible in this section; the other cells in the H position possess hairs that are located outside the field of view.
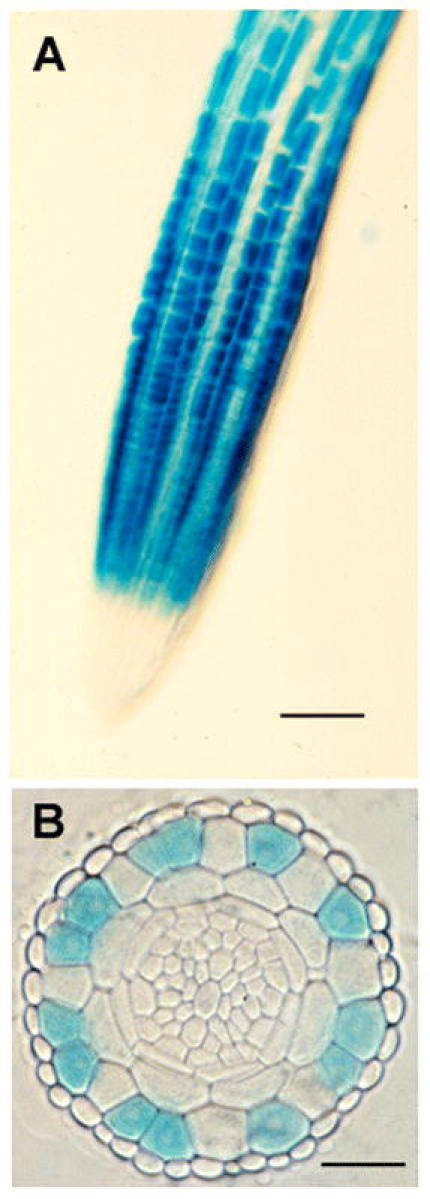
Expression of the GL2::GUS reporter fusion during root development. (A) Surface view showing preferential expression in the meristematic region. Bar = 50 µm. (B) Transverse section showing preferential expression in the N-cell position of the epidermis. Bar = 20 µm.
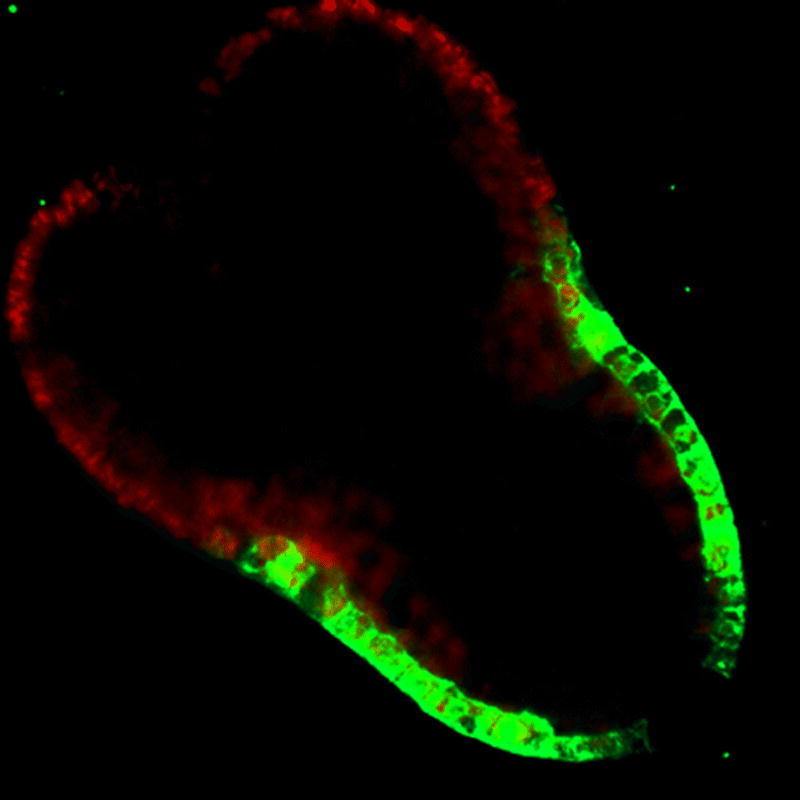
Embryonic expression of the GL2::GFP reporter fusion in the torpedo stage embryo. This median longitudinal view shows GFP accumulation in protodermal cells in the future hypocotyl and root.

Expression of the GL2::GUS reporter fusion in an epidermal cell clone derived from a rare longitudinal division. Note that only one set of cells in the clone expresses the GL2 marker. Bar = 10 µm.
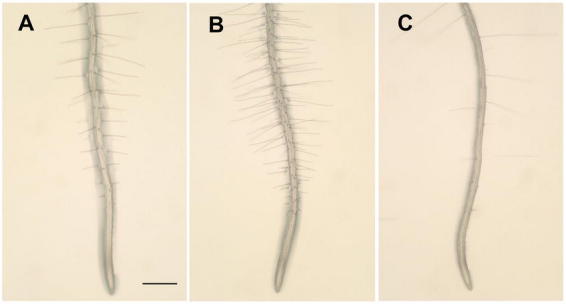
Root hair production in wild-type and cell specification mutants.(A) Wild-type.(B) An example of an ectopic hair mutant (wer).(C) An example of a reduced hair mutant (cpc).Bar = 500 µm for all images.
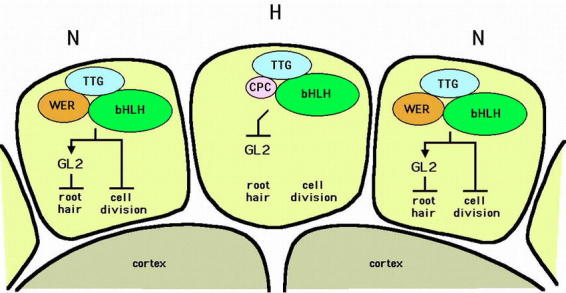
Model for the specification of the root hair and non-hair cell types in the Arabidopsis root epidermis. The proposed accumulation and interaction of cell fate regulators is shown within root epidermal cells destined to be root hair cells (in the H position) or non-hair cells (in the N position). In this model, the default fate for an epidermal cell is a root hair cell. Arrows indicated positive control, and blunted lines indicated negative regulation.
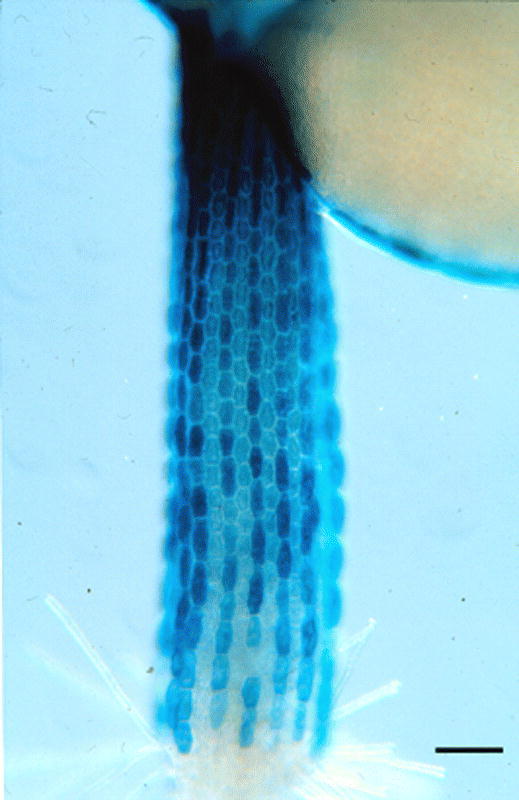
Expression of the GL2::GUS reporter fusion in the hypocotyl epidermis. Cells expressing the GL2::GUS marker are located in the N position. Bar = 100 µm.

Stages of root hair formation. Root hairs form in two main stages: initiation, when a small, disc-shaped area of the cell wall loosens to form a swelling, and tip growth, when the remainder of the hair grows by targeted secretion.
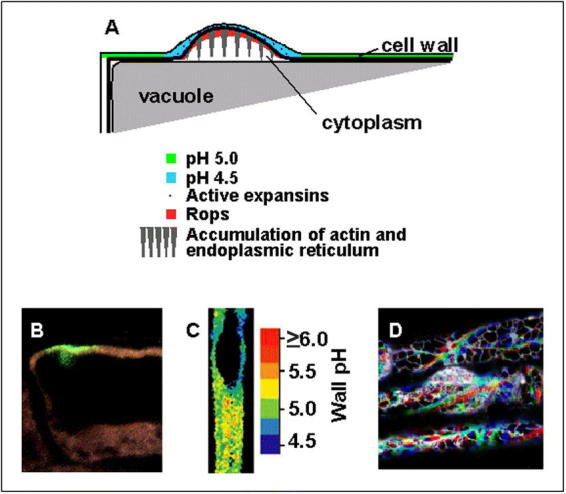
Root hair initiation.(A) Diagram summarizing the initiation process. Rop protein localizes to the initiation site, and the pH of the cell wall drops to about pH 4 to 4.5. This local pH change is thought to activate expansin proteins that loosen the cell wall. Large amounts of endoplasmic reticulum and filamentous (F) actin accumulate in the developing swelling(B) Rop at the future site of hair formation. Localization of Rop protein is the first sign that a hair is about to form (see Molendijk et al., 2001).(C) Acidification of the cell wall at the root hair initiation site. pH was imaged using NERF/Texas Red and pseudo-color coded according to the inset scale (see Bibikova et al., 1998).(D) Local accumulation of endoplasmic reticulum (ER) in an initiating hair flanked by two non-hair cells. Red, blue and green images were taken 30 seconds apart. White indicates that ER was present in the same location in all three images. (see Ridge et al., 1999).
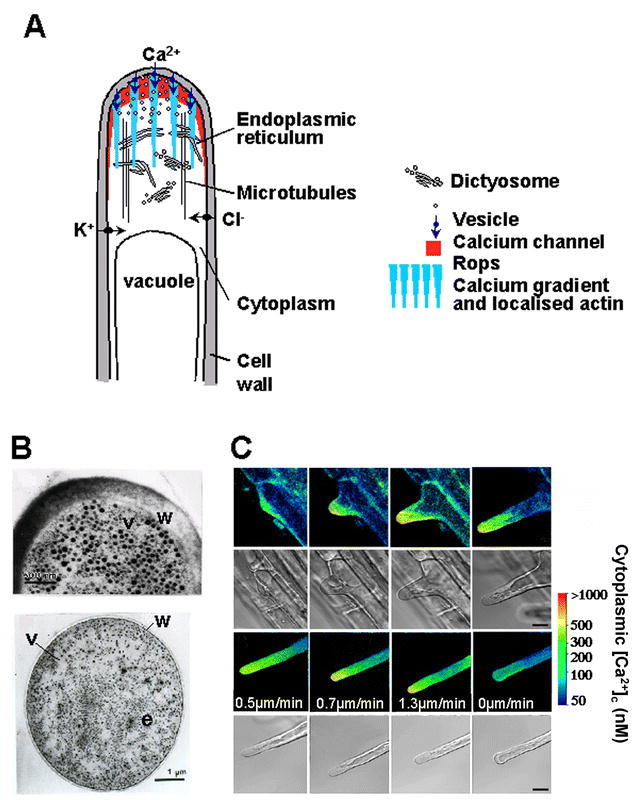
Root Hair Tip Growth.(A) Diagram summarizing the mechanism of tip growth in Arabidopsis root hairs. The tip is packed with membrane-bound vesicles delivering new cell wall material. These vesicles are made in the endoplasmic reticulum (ER) and dictyosomes which are abundant behind the tip. Rop protein is localized to the tip along with F-actin, and a tip-focused calcium gradient. This calcium gradient is thought to be generated by hyperpolarization-activated calcium channels, which are localized to the plasma membrane at the hair tip. Other channels import osmotically active K+ and Cl- ions, which help to sustain turgor pressure as the hair grows. The direction of growth is controlled by microtubules, which run along the length of the hair.(B) Cytoarchitecture at the tip of an elongating root hair. Transmission electron micrographs of sections of an elongating hair showing the cell wall (w), vesicles (v), and endoplasmic reticulum (e). Top – The hair apex is packed with vesicles. Bottom – A section from just behind the apex shows dense endoplasmic reticulum surrounded by vesicles.(C) Tip-growing root hairs have a tip-focused calcium gradient. Time course showing the establishment and maintenance of a calcium gradient in an elongating root hair, and its disappearance when growth ceases. The concentration of cytoplasmic free calcium ([Ca2+]c) was imaged using indo-1 and pseudo-color coded according to the inset scale. [Ca2+]c is shown in the first and third rows with corresponding transmitted light images of the same cell in the second and fourth rows (see Wymer et al. 1997).

A tip growing hair viewed with differential interference contrast microscopy (DIC). Most of the hair is vacuolated (V), there is an accumulation of cytoplasm at the tip (C), and the nucleus (N) (n denotes the nucleolus) has entered the hair.
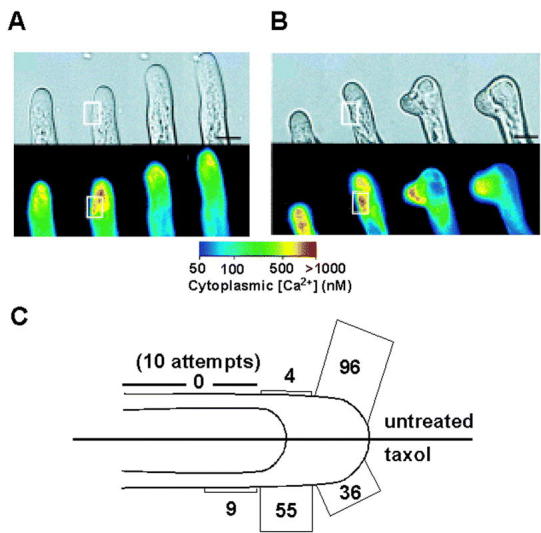
The direction of tip growth is controlled by calcium and microtubules.(A, B) Cytoplasmic calcium and tip growth in untreated (A) and 10µM taxol treated (B) root hairs after local photoactivation of caged calcium ionophore. The ionophore was activated by illuminating the boxed regions with a UV laser. The concentration of cytoplasmic free calcium ([Ca2+]c) was imaged using calcium green/rhodamine and pseudo-color coded according to the inset scale.(A) An untreated hair was unaffected by the ionophore.(B) A taxol-treated hair grew towards the ionophore. Taxol promotes microtubule polymerisation.(C) Diagram showing the percentage of untreated and taxol-treated root hairs that reoriented their growth in response to locally activated calcium ionophore (adapted from Bibikova et al. 1999).
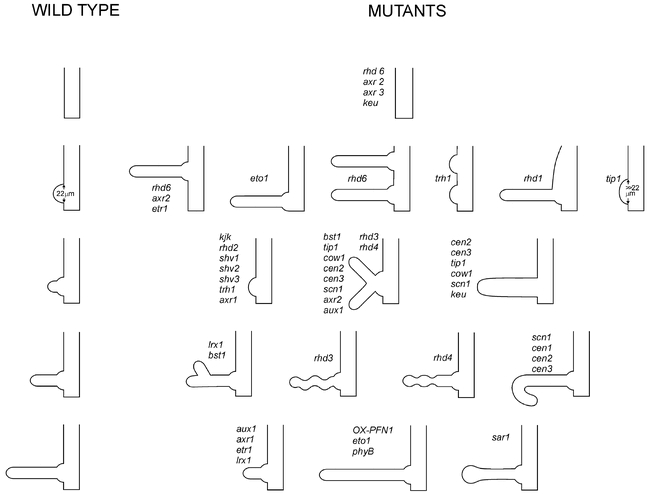
Genetics of root hair morphogenesis.Diagram summarizing the stages of root hair development that contribute to the shape of the hair cell, and the phenotypes of relevant mutants and transgenic plants. Root hairs are reduced in length to fit into the figure. The developmental stages of wild type hairs are shown on the left. The defects of mutant or transgenic hairs are shown on the right alongside the relevant stage of wild type development. Mutants appear more than once when they affect more than one stage of development. OX-PFN indicates over-expression of the PFN1 gene.
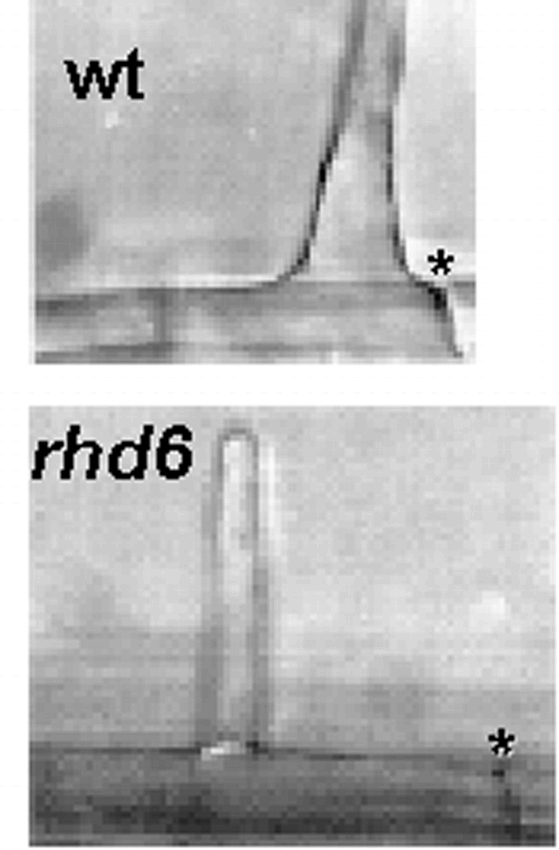
Hairs on the rhd6 mutant emerge at a more basal position on the hair cell than wild type hairs (wt). An asterisk (*) indicates the apical wall of each cell.
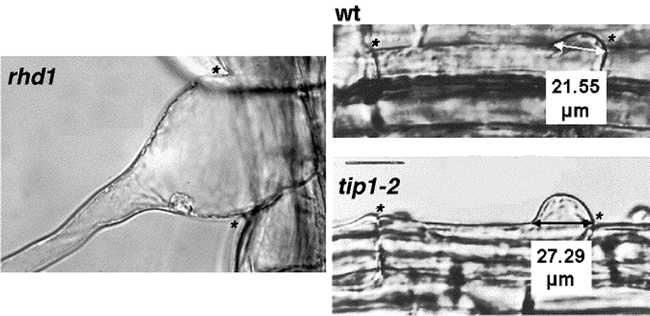
Swellings are wider on tip1 and rhd1 roots. Wild type swellings are about 22 µm across. Swellings on the rhd1 mutant encompass the whole outer cell wall. The tip1-2 mutant has swellings that are about 27 µm across. Asterisks (*) indicate the end walls of each cell (see Schiefelbein and Somerville, 1990; Parker et al., 2000).
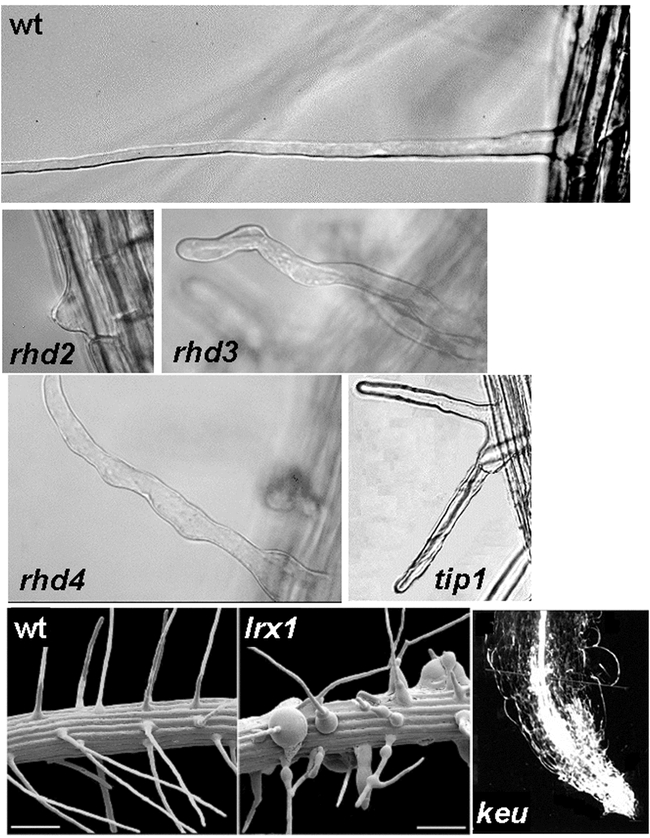
Phenotypes of wild type and mutant root hairs. Light micrographs of single hairs of wild type (wt), rhd2, rhd3, and rhd4, and a branched hair from a tip1 plant. Scanning electron micrographs of wild type (wt) and lrx1 roots. Light micrograph of a keule root tip (keu) with no root hairs. See Table 1 for references.
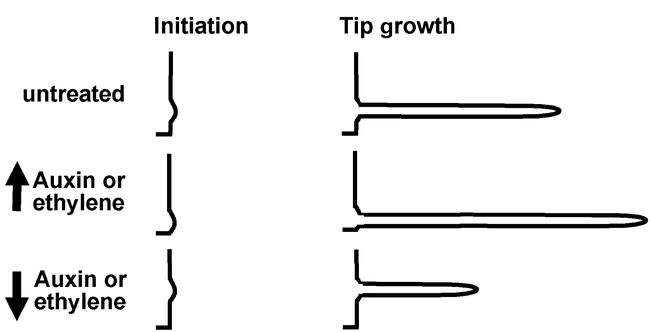
Roles of auxin and ethylene signaling in root hair growth.Increased auxin or ethylene signaling moves the intiation site to a more apical position and increases the amount of elongation during tip growth. Decreased auxin or ethylene signaling has the opposite effect.
Similar articles
-
The role of root hairs in water uptake: recent advances and future perspectives.J Exp Bot. 2022 Jun 2;73(11):3330-3338. doi: 10.1093/jxb/erac114. J Exp Bot. 2022. PMID: 35323893 Review.
-
The effect of root hairs on root water uptake is determined by root-soil contact and root hair shrinkage.New Phytol. 2023 Dec;240(6):2484-2497. doi: 10.1111/nph.19144. Epub 2023 Jul 31. New Phytol. 2023. PMID: 37525254
-
The root epidermis of Echium plantagineum L.: a novel type of pattern based on the distribution of short and long root hairs.Planta. 2003 Jun;217(2):238-44. doi: 10.1007/s00425-003-0989-4. Epub 2003 Feb 18. Planta. 2003. PMID: 12783331
-
PRX102 Participates in Root Hairs Tip Growth of Rice.Rice (N Y). 2023 Nov 16;16(1):51. doi: 10.1186/s12284-023-00668-7. Rice (N Y). 2023. PMID: 37971600 Free PMC article.
-
Root hairs vs. trichomes: Not everyone is straight!Curr Opin Plant Biol. 2021 Dec;64:102151. doi: 10.1016/j.pbi.2021.102151. Epub 2021 Dec 1. Curr Opin Plant Biol. 2021. PMID: 34864319 Review.
Cited by
-
Bacillus firmus I-1582 promotes plant growth and impairs infection and development of the cyst nematode Heterodera schachtii over two generations.Sci Rep. 2021 Jul 8;11(1):14114. doi: 10.1038/s41598-021-93567-0. Sci Rep. 2021. PMID: 34239009 Free PMC article.
-
Ectopic Expression of JcCPL1, 2, and 4 Affects Epidermal Cell Differentiation, Anthocyanin Biosynthesis and Leaf Senescence in Arabidopsis thaliana.Int J Mol Sci. 2022 Feb 9;23(4):1924. doi: 10.3390/ijms23041924. Int J Mol Sci. 2022. PMID: 35216041 Free PMC article.
-
Optimization of RNA In Situ Hybridization for mRNA Localization Detection in Mature Tissue of Cucumber Seedlings.Plants (Basel). 2020 Oct 29;9(11):1461. doi: 10.3390/plants9111461. Plants (Basel). 2020. PMID: 33138095 Free PMC article.
-
PFT1-controlled ROS balance is critical for multiple stages of root hair development in Arabidopsis.Plant Signal Behav. 2013 May;8(5):e24066. doi: 10.4161/psb.24066. Epub 2013 Mar 1. Plant Signal Behav. 2013. PMID: 23455023 Free PMC article.
-
VPS45 is required for both diffuse and tip growth of Arabidopsis thaliana cells.Front Plant Sci. 2023 Feb 27;14:1120307. doi: 10.3389/fpls.2023.1120307. eCollection 2023. Front Plant Sci. 2023. PMID: 36923123 Free PMC article.
References
-
- Baluska F., Salaj J., Mathur J., Braun M., Jasper F., Samaj J., Chua N. H., Barlow P. W., Volkmann D. Root hair formation: F-actin-dependent tip growth is initiated by local assembly of profilin-supported F-actin meshworks accumulated within expansin-enriched bulges. Dev. Biol. 2000;2271(1):618–632. - PubMed
-
- Bates T. R., Lynch J. P. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ. 1996;191(1):529–538.
-
- Bates T. R., Lynch J. P. Plant growth and phosphorus accumulation of wild type and two root hair mutants of Arabidopsis thaliana (Brassicaceae). Am. J. Bot. 2000a;871(1):958–963. - PubMed
-
- Bates T. R., Lynch J. P. The efficiency of Arabidopsis thaliana (Brassicaceae) root hairs in phosphorus acquisition. Am. J. Bot. 2000b;871(1):964–970. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources