

Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells
- PMID: 22279576
- PMCID: PMC3261192
- DOI: 10.1371/journal.pone.0030264
Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells
Abstract
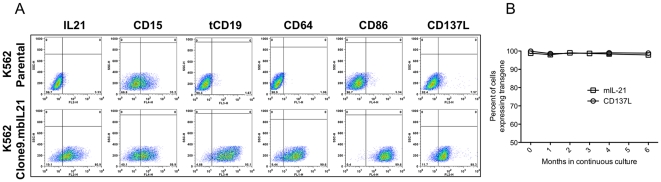
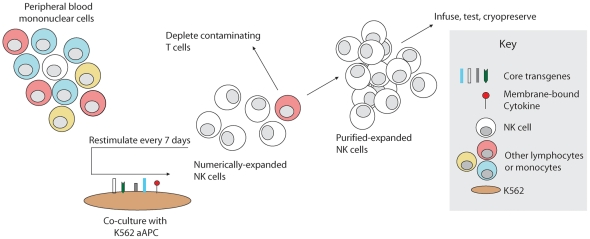
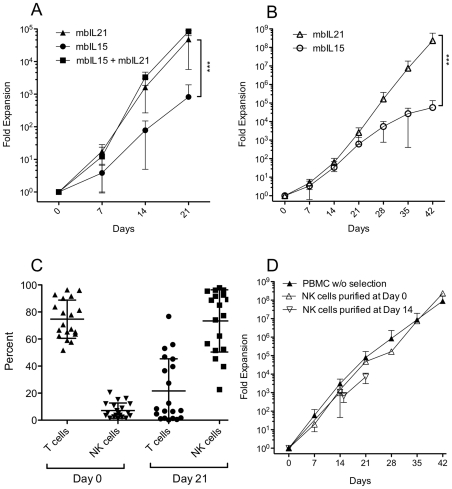
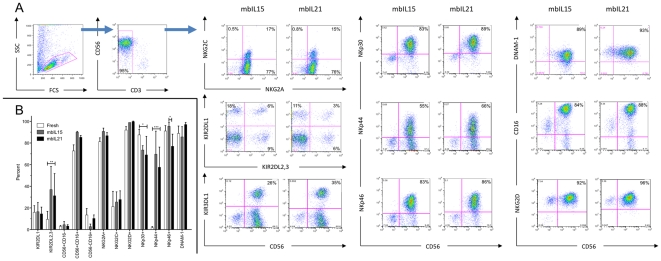
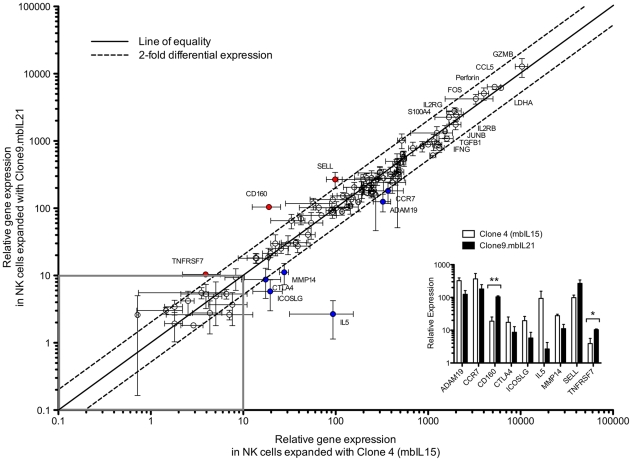
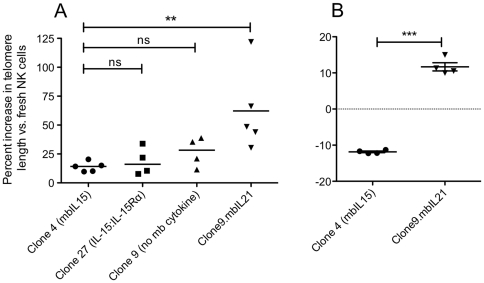
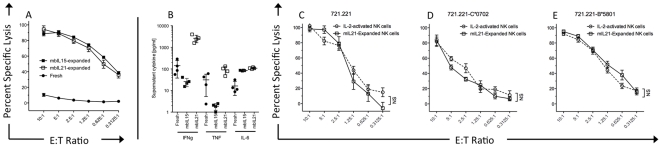
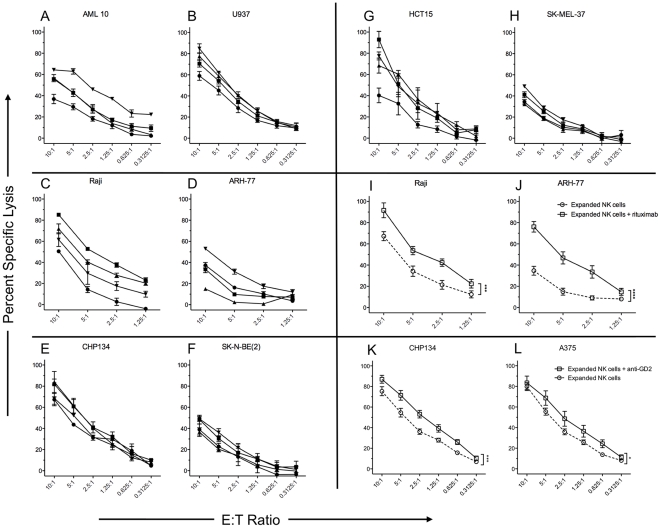
NK cells have therapeutic potential for a wide variety of human malignancies. However, because NK cells expand poorly in vitro, have limited life spans in vivo, and represent a small fraction of peripheral white blood cells, obtaining sufficient cell numbers is the major obstacle for NK-cell immunotherapy. Genetically-engineered artificial antigen-presenting cells (aAPCs) expressing membrane-bound IL-15 (mbIL15) have been used to propagate clinical-grade NK cells for human trials of adoptive immunotherapy, but ex vivo proliferation has been limited by telomere shortening. We developed K562-based aAPCs with membrane-bound IL-21 (mbIL21) and assessed their ability to support human NK-cell proliferation. In contrast to mbIL15, mbIL21-expressing aAPCs promoted log-phase NK cell expansion without evidence of senescence for up to 6 weeks of culture. By day 21, parallel expansion of NK cells from 22 donors demonstrated a mean 47,967-fold expansion (median 31,747) when co-cultured with aAPCs expressing mbIL21 compared to 825-fold expansion (median 325) with mbIL15. Despite the significant increase in proliferation, mbIL21-expanded NK cells also showed a significant increase in telomere length compared to freshly obtained NK cells, suggesting a possible mechanism for their sustained proliferation. NK cells expanded with mbIL21 were similar in phenotype and cytotoxicity to those expanded with mbIL15, with retained donor KIR repertoires and high expression of NCRs, CD16, and NKG2D, but had superior cytokine secretion. The mbIL21-expanded NK cells showed increased transcription of the activating receptor CD160, but otherwise had remarkably similar mRNA expression profiles of the 96 genes assessed. mbIL21-expanded NK cells had significant cytotoxicity against all tumor cell lines tested, retained responsiveness to inhibitory KIR ligands, and demonstrated enhanced killing via antibody-dependent cell cytotoxicity. Thus, aAPCs expressing mbIL21 promote improved proliferation of human NK cells with longer telomeres and less senescence, supporting their clinical use in propagating NK cells for adoptive immunotherapy.
Conflict of interest statement
Figures
Similar articles
-
Autonomous growth and increased cytotoxicity of natural killer cells expressing membrane-bound interleukin-15.Blood. 2014 Aug 14;124(7):1081-8. doi: 10.1182/blood-2014-02-556837. Epub 2014 Jul 8. Blood. 2014. PMID: 25006133
-
Sequential Exposure to IL21 and IL15 During Human Natural Killer Cell Expansion Optimizes Yield and Function.Cancer Immunol Res. 2023 Nov 1;11(11):1524-1537. doi: 10.1158/2326-6066.CIR-23-0151. Cancer Immunol Res. 2023. PMID: 37649085 Free PMC article.
-
Ex Vivo Expansion of Human NK Cells Using K562 Engineered to Express Membrane Bound IL21.Methods Mol Biol. 2016;1441:175-93. doi: 10.1007/978-1-4939-3684-7_15. Methods Mol Biol. 2016. PMID: 27177666
-
Selection and expansion of natural killer cells for NK cell-based immunotherapy.Cancer Immunol Immunother. 2016 Apr;65(4):477-84. doi: 10.1007/s00262-016-1792-y. Epub 2016 Jan 25. Cancer Immunol Immunother. 2016. PMID: 26810567 Free PMC article. Review.
-
Cellular therapy: Adoptive immunotherapy with expanded natural killer cells.Immunol Rev. 2019 Jul;290(1):85-99. doi: 10.1111/imr.12793. Immunol Rev. 2019. PMID: 31355489 Review.
Cited by
-
Clinical-scale derivation of natural killer cells from human pluripotent stem cells for cancer therapy.Stem Cells Transl Med. 2013 Apr;2(4):274-83. doi: 10.5966/sctm.2012-0084. Epub 2013 Mar 20. Stem Cells Transl Med. 2013. PMID: 23515118 Free PMC article.
-
Engineered human embryonic stem cell-derived lymphocytes to study in vivo trafficking and immunotherapy.Stem Cells Dev. 2013 Jul 1;22(13):1861-9. doi: 10.1089/scd.2012.0608. Epub 2013 Mar 28. Stem Cells Dev. 2013. PMID: 23421330 Free PMC article.
-
Harnessing natural killer cells for refractory/relapsed non-Hodgkin lymphoma: biological roles, clinical trials, and future prospective.Biomark Res. 2024 Jul 17;12(1):66. doi: 10.1186/s40364-024-00610-z. Biomark Res. 2024. PMID: 39020411 Free PMC article. Review.
-
"Adherent" versus Other Isolation Strategies for Expanding Purified, Potent, and Activated Human NK Cells for Cancer Immunotherapy.Biomed Res Int. 2015;2015:869547. doi: 10.1155/2015/869547. Epub 2015 Jun 16. Biomed Res Int. 2015. PMID: 26161419 Free PMC article. Review.
-
Cellular Strategies for Separating GvHD from GvL in Haploidentical Transplantation.Cells. 2024 Jan 11;13(2):134. doi: 10.3390/cells13020134. Cells. 2024. PMID: 38247827 Free PMC article. Review.
References
-
- Barao I, Murphy WJ. The immunobiology of natural killer cells and bone marrow allograft rejection. Biol Blood Marrow Transplant. 2003;9:727–741. - PubMed
-
- Robertson MJ, Ritz J. Biology and clinical relevance of human natural killer cells. Blood. 1990;76:2421–2438. - PubMed
-
- Farag SS, Fehniger TA, Ruggeri L, Velardi A, Caligiuri MA. Natural killer cell receptors: new biology and insights into the graft-versus-leukemia effect. Blood. 2002;100:1935–1947. - PubMed
-
- Herberman RB, Ortaldo JR. Natural killer cells: their roles in defenses against disease. Science. 1981;214:24–30. - PubMed
-
- Pittari G, Fregni G, Roguet L, Garcia A, Vataire AL, et al. Early evaluation of natural killer activity in post-transplant acute myeloid leukemia patients. Bone Marrow Transplant. 2010;45:862–871. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
