

Activation of legumain involves proteolytic and conformational events, resulting in a context- and substrate-dependent activity profile
- PMID: 22232165
- PMCID: PMC3253828
- DOI: 10.1107/S1744309111048020
Activation of legumain involves proteolytic and conformational events, resulting in a context- and substrate-dependent activity profile
Abstract
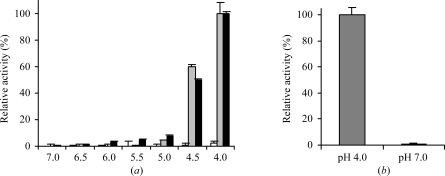
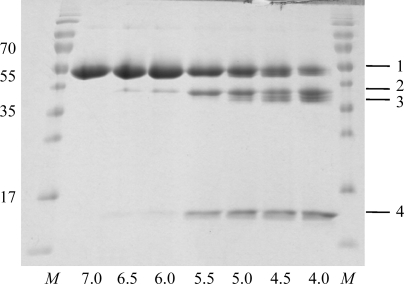
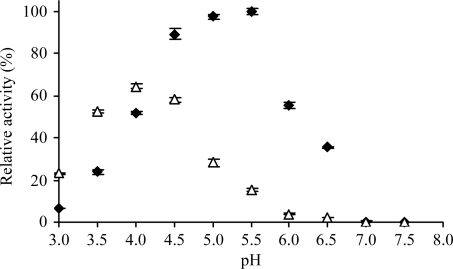
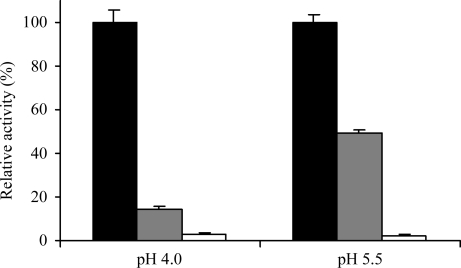
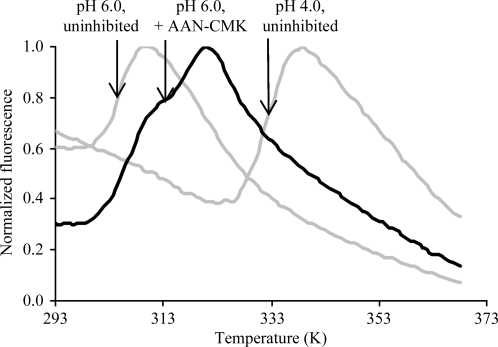
Localized mainly to endo/lysosomes, legumain plays an important role in exogenous antigen processing and presentation. The cysteine protease legumain, also known as asparaginyl endopepetidase AEP, is synthesized as a zymogen and is known to undergo pH-dependent autoproteolytic activation whereby N-terminal and C-terminal propeptides are released. However, important mechanistic details of this pH-dependent activation as well as the characteristic pH activity profile remain unclear. Here, it is shown that all but one of the autocatalytic cleavage events occur in trans, with only the release of the C-terminal propeptide being relevant to enzymatic activity. An intriguing super-activation event that appears to be exclusively conformational in nature and enhances the enzymatic activity of proteolytically fully processed legumain by about twofold was also found. Accepting asparagines and, to lesser extent, aspartic acid in P1, super-activated legumain exhibits a marked pH dependence that is governed by the P1 residue of its substrate and conformationally stabilizing factors such as temperature or ligands. The crystallization and preliminary diffraction data analysis of active legumain are presented, which form an important basis for further studies that should clarify fundamental aspects of activation, activity and inactivation of legumain, which is a key target in (auto-)immunity and cancer.
© 2012 International Union of Crystallography. All rights reserved.
Figures


Similar articles
-
Mechanistic and structural studies on legumain explain its zymogenicity, distinct activation pathways, and regulation.Proc Natl Acad Sci U S A. 2013 Jul 2;110(27):10940-5. doi: 10.1073/pnas.1300686110. Epub 2013 Jun 17. Proc Natl Acad Sci U S A. 2013. PMID: 23776206 Free PMC article.
-
Structure and function of legumain in health and disease.Biochimie. 2016 Mar;122:126-50. doi: 10.1016/j.biochi.2015.09.022. Epub 2015 Sep 25. Biochimie. 2016. PMID: 26403494 Review.
-
Structural analysis of asparaginyl endopeptidase reveals the activation mechanism and a reversible intermediate maturation stage.Cell Res. 2014 Mar;24(3):344-58. doi: 10.1038/cr.2014.4. Epub 2014 Jan 10. Cell Res. 2014. PMID: 24407422 Free PMC article.
-
Synthesis and evaluation of aza-peptidyl inhibitors of the lysosomal asparaginyl endopeptidase, legumain.Bioorg Med Chem Lett. 2012 Feb 1;22(3):1340-3. doi: 10.1016/j.bmcl.2011.12.079. Epub 2011 Dec 21. Bioorg Med Chem Lett. 2012. PMID: 22243962 Free PMC article.
-
The Mammalian Cysteine Protease Legumain in Health and Disease.Int J Mol Sci. 2022 Dec 15;23(24):15983. doi: 10.3390/ijms232415983. Int J Mol Sci. 2022. PMID: 36555634 Free PMC article. Review.
Cited by
-
The TvLEGU-1, a legumain-like cysteine proteinase, plays a key role in Trichomonas vaginalis cytoadherence.Biomed Res Int. 2013;2013:561979. doi: 10.1155/2013/561979. Epub 2013 Jan 1. Biomed Res Int. 2013. PMID: 23509742 Free PMC article.
-
Legumain is a paracrine regulator of osteoblast differentiation and mediates the inhibitory effect of TGF-β1 on osteoblast maturation.Front Endocrinol (Lausanne). 2024 Sep 19;15:1445049. doi: 10.3389/fendo.2024.1445049. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 39363898 Free PMC article.
-
Chemically Programmed Bispecific Antibody Targeting Legumain Protease and αvβ3 Integrin Mediates Strong Antitumor Effects.Mol Pharm. 2015 Jul 6;12(7):2544-50. doi: 10.1021/acs.molpharmaceut.5b00257. Epub 2015 Jun 9. Mol Pharm. 2015. PMID: 26024761 Free PMC article.
-
The legumain McPAL1 from Momordica cochinchinensis is a highly stable Asx-specific splicing enzyme.J Biol Chem. 2021 Dec;297(6):101325. doi: 10.1016/j.jbc.2021.101325. Epub 2021 Oct 26. J Biol Chem. 2021. PMID: 34710371 Free PMC article.
-
Crystal Structure of Plant Legumain Reveals a Unique Two-Chain State with pH-Dependent Activity Regulation.Plant Cell. 2018 Mar;30(3):686-699. doi: 10.1105/tpc.17.00963. Epub 2018 Feb 16. Plant Cell. 2018. PMID: 29453229 Free PMC article.
References
-
- Adams, P. D., Grosse-Kunstleve, R. W., Hung, L.-W., Ioerger, T. R., McCoy, A. J., Moriarty, N. W., Read, R. J., Sacchettini, J. C., Sauter, N. K. & Terwilliger, T. C. (2002). Acta Cryst. D58, 1948–1954. - PubMed
-
- Alim, M. A., Tsuji, N., Miyoshi, T., Islam, M. K., Huang, X., Hatta, T. & Fujisaki, K. (2008). J. Insect Physiol. 54, 573–585. - PubMed
-
- Alvarez-Fernandez, M., Barrett, A. J., Gerhartz, B., Dando, P. M., Ni, J. & Abrahamson, M. (1999). J. Biol. Chem. 274, 19195–19203. - PubMed
-
- Baker, D., Shiau, A. K. & Agard, D. A. (1993). Curr. Opin. Cell Biol. 5, 966–970. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
