

Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors
- PMID: 22198272
- PMCID: PMC3271782
- DOI: 10.1104/pp.111.188623
Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors
Abstract
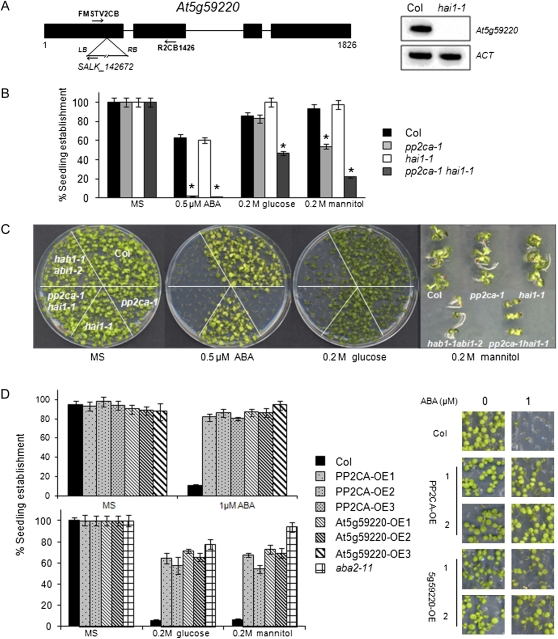
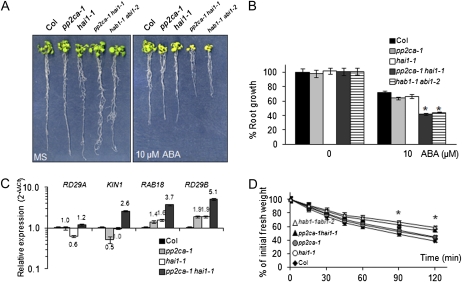
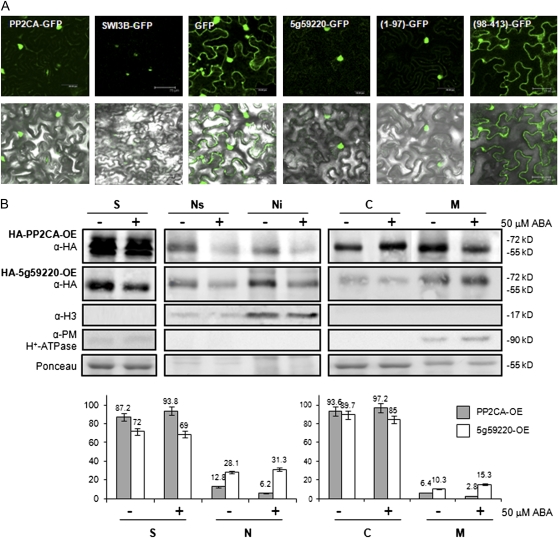
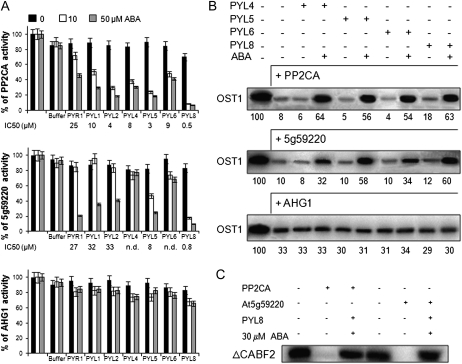
Clade A protein phosphatases type 2C (PP2Cs) are negative regulators of abscisic acid (ABA) signaling that are inhibited in an ABA-dependent manner by PYRABACTIN RESISTANCE1 (PYR1)/PYR1-LIKE (PYL)/REGULATORY COMPONENTS OF ABA RECEPTORS (RCAR) intracellular receptors. We provide genetic evidence that a previously uncharacterized member of this PP2C family in Arabidopsis (Arabidopsis thaliana), At5g59220, is a negative regulator of osmotic stress and ABA signaling and that this function was only apparent when double loss-of-function mutants with pp2ca-1/ahg3 were generated. At5g59220-green fluorescent protein and its close relative PP2CA-green fluorescent protein showed a predominant nuclear localization; however, hemagglutinin-tagged versions were also localized to cytosol and microsomal pellets. At5g59220 was selectively inhibited by some PYR/PYL ABA receptors, and close relatives of this PP2C, such as PP2CA/ABA-HYPERSENSITIVE GERMINATION3 (AHG3) and AHG1, showed a contrasting sensitivity to PYR/PYL inhibition. Interestingly, AHG1 was resistant to inhibition by the PYR/PYL receptors tested, which suggests that this seed-specific phosphatase is still able to regulate ABA signaling in the presence of ABA and PYR/PYL receptors and therefore to control the highly active ABA signaling pathway that operates during seed development. Moreover, the differential sensitivity of the phosphatases At5g59220 and PP2CA to inhibition by ABA receptors reveals a functional specialization of PYR/PYL ABA receptors to preferentially inhibit certain PP2Cs.
Figures
Similar articles
-
Tomato PYR/PYL/RCAR abscisic acid receptors show high expression in root, differential sensitivity to the abscisic acid agonist quinabactin, and the capability to enhance plant drought resistance.J Exp Bot. 2014 Aug;65(15):4451-64. doi: 10.1093/jxb/eru219. Epub 2014 May 26. J Exp Bot. 2014. PMID: 24863435 Free PMC article.
-
PYRABACTIN RESISTANCE1-LIKE8 plays an important role for the regulation of abscisic acid signaling in root.Plant Physiol. 2013 Feb;161(2):931-41. doi: 10.1104/pp.112.208678. Epub 2012 Dec 14. Plant Physiol. 2013. PMID: 23370718 Free PMC article.
-
Inactivation of PYR/PYL/RCAR ABA receptors by tyrosine nitration may enable rapid inhibition of ABA signaling by nitric oxide in plants.Sci Signal. 2015 Sep 1;8(392):ra89. doi: 10.1126/scisignal.aaa7981. Sci Signal. 2015. PMID: 26329583
-
Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs.Plant Sci. 2012 Jan;182:3-11. doi: 10.1016/j.plantsci.2010.11.014. Epub 2010 Dec 7. Plant Sci. 2012. PMID: 22118610 Review.
-
[ABA signaling mediated by PYR/PYL/RCAR in plants].Yi Chuan. 2012 May;34(5):560-72. doi: 10.3724/sp.j.1005.2012.00560. Yi Chuan. 2012. PMID: 22659428 Review. Chinese.
Cited by
-
Type 2C phosphatase 1 of Artemisia annua L. is a negative regulator of ABA signaling.Biomed Res Int. 2014;2014:521794. doi: 10.1155/2014/521794. Epub 2014 Oct 28. Biomed Res Int. 2014. PMID: 25530962 Free PMC article.
-
TaPYL4, an ABA receptor gene of wheat, positively regulates plant drought adaptation through modulating the osmotic stress-associated processes.BMC Plant Biol. 2022 Sep 1;22(1):423. doi: 10.1186/s12870-022-03799-z. BMC Plant Biol. 2022. PMID: 36050643 Free PMC article.
-
The functional analysis of SlNCED1 in tomato pollen development.Cell Mol Life Sci. 2018 Sep;75(18):3457-3472. doi: 10.1007/s00018-018-2809-9. Epub 2018 Apr 9. Cell Mol Life Sci. 2018. PMID: 29632966 Free PMC article.
-
Related to ABA-Insensitive3(ABI3)/Viviparous1 and AtABI5 transcription factor coexpression in cotton enhances drought stress adaptation.Plant Biotechnol J. 2014 Jun;12(5):578-89. doi: 10.1111/pbi.12162. Epub 2014 Feb 1. Plant Biotechnol J. 2014. PMID: 24483851 Free PMC article.
-
Arabidopsis Duodecuple Mutant of PYL ABA Receptors Reveals PYL Repression of ABA-Independent SnRK2 Activity.Cell Rep. 2018 Jun 12;23(11):3340-3351.e5. doi: 10.1016/j.celrep.2018.05.044. Cell Rep. 2018. PMID: 29898403 Free PMC article.
References
-
- Baykov AA, Evtushenko OA, Avaeva SM. (1988) A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Anal Biochem 171: 266–270 - PubMed
-
- Bensmihen S, To A, Lambert G, Kroj T, Giraudat J, Parcy F. (2004) Analysis of an activated ABI5 allele using a new selection method for transgenic Arabidopsis seeds. FEBS Lett 561: 127–131 - PubMed
-
- Bowler C, Benvenuto G, Laflamme P, Molino D, Probst AV, Tariq M, Paszkowski J. (2004) Chromatin techniques for plant cells. Plant J 39: 776–789 - PubMed
-
- Cho YH, Yoo SD, Sheen J. (2006) Regulatory functions of nuclear hexokinase1 complex in glucose signaling. Cell 127: 579–589 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
