

Intracellular neutralization of viral infection in polarized epithelial cells by neonatal Fc receptor (FcRn)-mediated IgG transport
- PMID: 22042859
- PMCID: PMC3215070
- DOI: 10.1073/pnas.1115348108
Intracellular neutralization of viral infection in polarized epithelial cells by neonatal Fc receptor (FcRn)-mediated IgG transport
Abstract
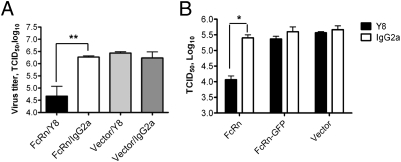
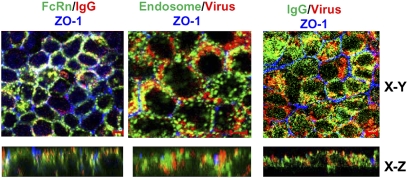
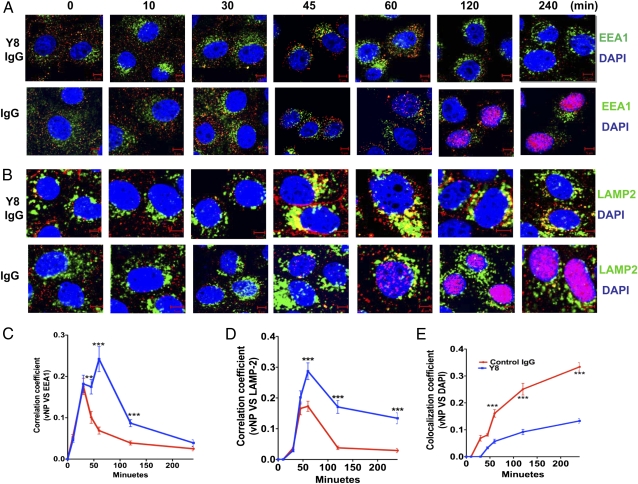
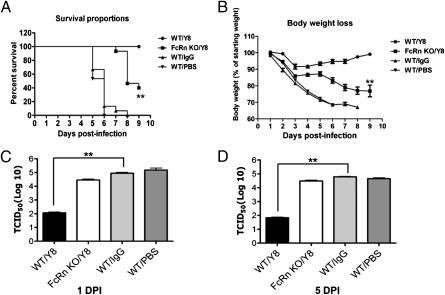
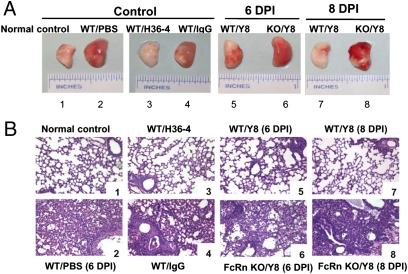
IgG was traditionally thought to neutralize virions by blocking their attachment to or penetration into mucosal epithelial cells, a common site of exposure to viruses. However, we describe an intracellular neutralizing action for an influenza hemagglutinin-specific monoclonal antibody, Y8-10C2 (Y8), which has neutralizing activity only at an acidic pH. When Y8 was applied to the basolateral surface of Madin-Darby canine kidney cells expressing the rat neonatal Fc receptor for IgG (FcRn), it significantly reduced viral replication following apical exposure of the cell monolayer to influenza virus. Virus neutralization by Y8 mAb was dependent on FcRn expression and its transport of IgG. As both FcRn and Y8 mAb bind their partners only at acidic pH, the Y8 mAb is proposed to carry out its antiviral activity intracellularly. Furthermore, the virus, Y8 mAb, and FcRn colocalized within endosomes, possibly inhibiting the fusion of viral envelopes with endosomal membranes during primary uncoating, and preventing the accumulation of the neutralized viral nucleoprotein antigen in the nucleus. Prophylactic administration of Y8 mAb before viral challenge in WT mice, but not FcRn-KO mice, conferred protection from lethality, prevented weight loss, resulted in a significant reduction in pulmonary virus titers, and largely reduced virus-induced lung pathology. Thus, this study reveals an intracellular mechanism for viral neutralization in polarized epithelial cells that is dependent on FcRn-mediated transport of neutralizing IgG.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
FcRn-Targeted Mucosal Vaccination against Influenza Virus Infection.J Immunol. 2021 Sep 1;207(5):1310-1321. doi: 10.4049/jimmunol.2100297. Epub 2021 Aug 11. J Immunol. 2021. PMID: 34380652 Free PMC article.
-
Roles of Fc Domain and Exudation in L2 Antibody-Mediated Protection against Human Papillomavirus.J Virol. 2018 Jul 17;92(15):e00572-18. doi: 10.1128/JVI.00572-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29743371 Free PMC article.
-
Bidirectional transcytosis of IgG by the rat neonatal Fc receptor expressed in a rat kidney cell line: a system to study protein transport across epithelia.J Cell Sci. 2000 Apr;113 ( Pt 7):1277-85. doi: 10.1242/jcs.113.7.1277. J Cell Sci. 2000. PMID: 10704378
-
Are endosomal trafficking parameters better targets for improving mAb pharmacokinetics than FcRn binding affinity?Mol Immunol. 2013 Dec;56(4):660-74. doi: 10.1016/j.molimm.2013.05.008. Epub 2013 Aug 2. Mol Immunol. 2013. PMID: 23917469 Review.
-
New functions of the MHC class I-related Fc receptor, FcRn.Biochem Soc Trans. 1997 May;25(2):481-6. doi: 10.1042/bst0250481. Biochem Soc Trans. 1997. PMID: 9191140 Review.
Cited by
-
Extra-Neutralizing FcR-Mediated Antibody Functions for a Universal Influenza Vaccine.Front Immunol. 2019 Mar 18;10:440. doi: 10.3389/fimmu.2019.00440. eCollection 2019. Front Immunol. 2019. PMID: 30949165 Free PMC article. Review.
-
FcRn: The Architect Behind the Immune and Nonimmune Functions of IgG and Albumin.J Immunol. 2015 May 15;194(10):4595-603. doi: 10.4049/jimmunol.1403014. J Immunol. 2015. PMID: 25934922 Free PMC article. Review.
-
Intracellular antibody immunity.J Clin Immunol. 2014 Jul;34 Suppl 1:S30-4. doi: 10.1007/s10875-014-0017-4. Epub 2014 Apr 11. J Clin Immunol. 2014. PMID: 24722852 Review.
-
TRIM21: a cytosolic Fc receptor with broad antibody isotype specificity.Immunol Rev. 2015 Nov;268(1):328-39. doi: 10.1111/imr.12363. Immunol Rev. 2015. PMID: 26497531 Free PMC article. Review.
-
Plasma cell and serum antibody responses to influenza vaccine in preterm and full-term infants.Vaccine. 2017 Sep 12;35(38):5163-5171. doi: 10.1016/j.vaccine.2017.07.115. Epub 2017 Aug 12. Vaccine. 2017. PMID: 28807607 Free PMC article.
References
-
- Roopenian DC, Akilesh S. FcRn: The neonatal Fc receptor comes of age. Nat Rev Immunol. 2007;7:715–725. - PubMed
-
- Raghavan M, Bonagura VR, Morrison SL, Bjorkman PJ. Analysis of the pH dependence of the neonatal Fc receptor/immunoglobulin G interaction using antibody and receptor variants. Biochemistry. 1995;34:14649–14657. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
