

The cytomegaloviral protein pUL138 acts as potentiator of tumor necrosis factor (TNF) receptor 1 surface density to enhance ULb'-encoded modulation of TNF-α signaling
- PMID: 21976655
- PMCID: PMC3233134
- DOI: 10.1128/JVI.06005-11
The cytomegaloviral protein pUL138 acts as potentiator of tumor necrosis factor (TNF) receptor 1 surface density to enhance ULb'-encoded modulation of TNF-α signaling
Abstract
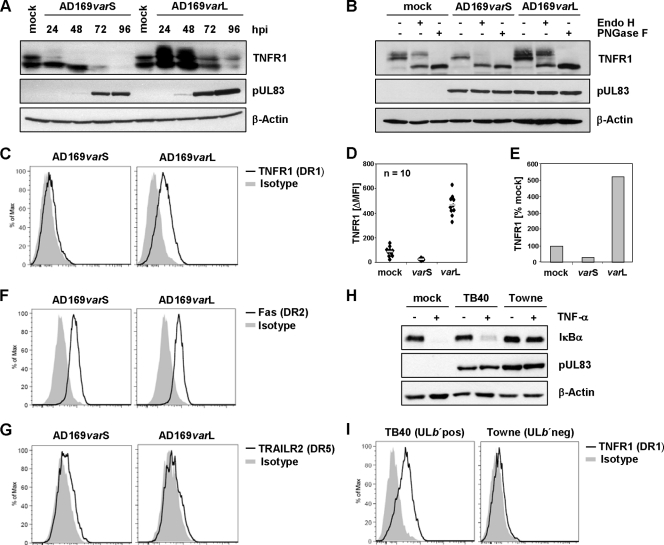
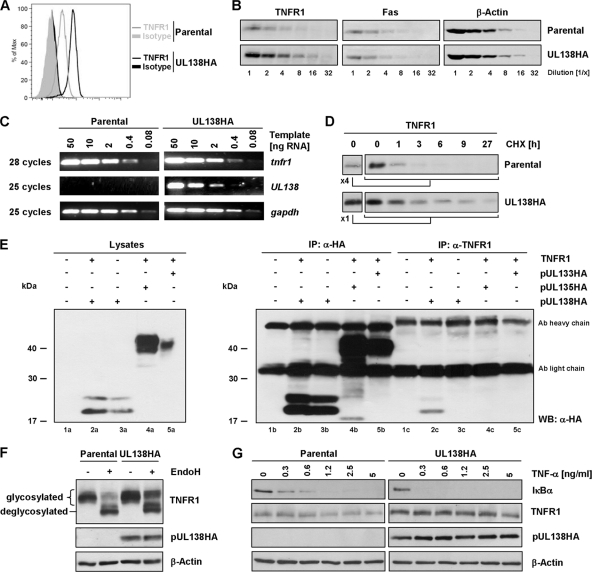
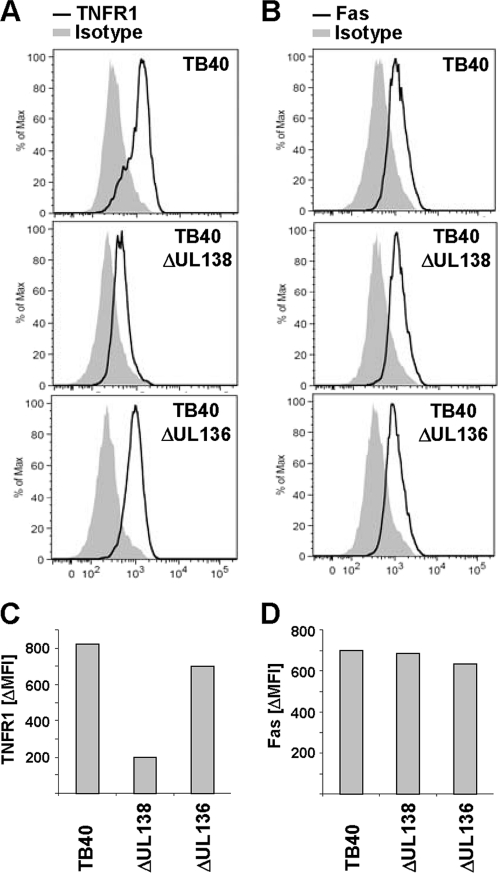
Human cytomegalovirus is a ubiquitous herpesvirus that establishes lifelong latent infection. Changes in immune homeostasis induce the reactivation of lytic infection, which is mostly inapparent in healthy individuals but often causes overt disease in immunocompromised hosts. Based on discrepant tumor necrosis factor receptor 1 surface disposition between human cytomegalovirus AD169 variants differing in the ULb' region, we identified the latency-associated gene product pUL138, which also is expressed during productive infection, as a selective potentiator of tumor necrosis factor receptor 1, one of the key receptors of innate immunity. Ectopically expressed pUL138 coprecipitated with tumor necrosis factor receptor 1, extended the protein half-life, and enhanced its signaling responses, thus leading to tumor necrosis factor receptor 1 hyperresponsiveness. Conversely, the targeted deletion of UL138 from the human cytomegaloviral genome strongly reduced tumor necrosis factor receptor 1 surface densities of infected cells. Remarkably, the comparison of UL138 deficiency to ULb' deficiency revealed the presence of further positive modulators of tumor necrosis factor alpha signal transduction encoded within the human cytomegalovirus ULb' region, identifying this region as a hub for multilayered tumor necrosis factor alpha signaling regulation.
Figures



Similar articles
-
The latency-associated UL138 gene product of human cytomegalovirus sensitizes cells to tumor necrosis factor alpha (TNF-alpha) signaling by upregulating TNF-alpha receptor 1 cell surface expression.J Virol. 2011 Nov;85(21):11409-21. doi: 10.1128/JVI.05028-11. Epub 2011 Aug 31. J Virol. 2011. PMID: 21880774 Free PMC article.
-
A novel human cytomegalovirus locus modulates cell type-specific outcomes of infection.PLoS Pathog. 2011 Dec;7(12):e1002444. doi: 10.1371/journal.ppat.1002444. Epub 2011 Dec 29. PLoS Pathog. 2011. PMID: 22241980 Free PMC article.
-
Interactions between proteins encoded within the human cytomegalovirus UL133-UL138 locus.J Virol. 2012 Aug;86(16):8653-62. doi: 10.1128/JVI.00465-12. Epub 2012 Jun 6. J Virol. 2012. PMID: 22674978 Free PMC article.
-
Deletion of the non-adjacent genes UL148 and UL148D impairs human cytomegalovirus-mediated TNF receptor 2 surface upregulation.Front Immunol. 2023 Aug 3;14:1170300. doi: 10.3389/fimmu.2023.1170300. eCollection 2023. Front Immunol. 2023. PMID: 37600801 Free PMC article.
-
Role of TNF-TNF Receptor 2 Signal in Regulatory T Cells and Its Therapeutic Implications.Front Immunol. 2018 Apr 19;9:784. doi: 10.3389/fimmu.2018.00784. eCollection 2018. Front Immunol. 2018. PMID: 29725328 Free PMC article. Review.
Cited by
-
Human cytomegalovirus Fcγ binding proteins gp34 and gp68 antagonize Fcγ receptors I, II and III.PLoS Pathog. 2014 May 15;10(5):e1004131. doi: 10.1371/journal.ppat.1004131. eCollection 2014 May. PLoS Pathog. 2014. PMID: 24830376 Free PMC article.
-
Multimodal HLA-I genotype regulation by human cytomegalovirus US10 and resulting surface patterning.Elife. 2024 Jun 20;13:e85560. doi: 10.7554/eLife.85560. Elife. 2024. PMID: 38900146 Free PMC article.
-
Die Another Day: Inhibition of Cell Death Pathways by Cytomegalovirus.Viruses. 2017 Sep 2;9(9):249. doi: 10.3390/v9090249. Viruses. 2017. PMID: 28869497 Free PMC article. Review.
-
Human cytomegalovirus antagonizes activation of Fcγ receptors by distinct and synergizing modes of IgG manipulation.Elife. 2021 Mar 16;10:e63877. doi: 10.7554/eLife.63877. Elife. 2021. PMID: 33724188 Free PMC article.
-
The human cytomegalovirus UL133-138 gene locus attenuates the lytic viral cycle in fibroblasts.PLoS One. 2015 Mar 23;10(3):e0120946. doi: 10.1371/journal.pone.0120946. eCollection 2015. PLoS One. 2015. PMID: 25799165 Free PMC article.
References
-
- Benedict C. A., et al. 2001. Three adenovirus E3 proteins cooperate to evade apoptosis by tumor necrosis factor-related apoptosis-inducing ligand receptor-1 and -2. J. Biol. Chem. 276:3270–3278 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
