

Cooperative roles of fish protein kinase containing Z-DNA binding domains and double-stranded RNA-dependent protein kinase in interferon-mediated antiviral response
- PMID: 21937641
- PMCID: PMC3209354
- DOI: 10.1128/JVI.05849-11
Cooperative roles of fish protein kinase containing Z-DNA binding domains and double-stranded RNA-dependent protein kinase in interferon-mediated antiviral response
Abstract
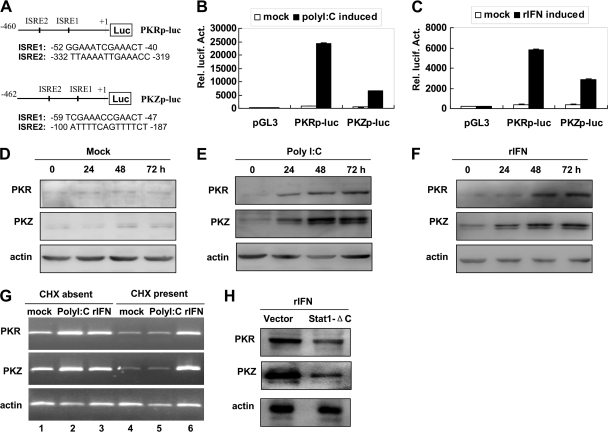
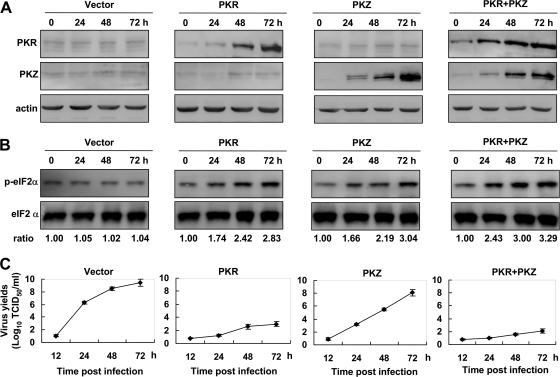
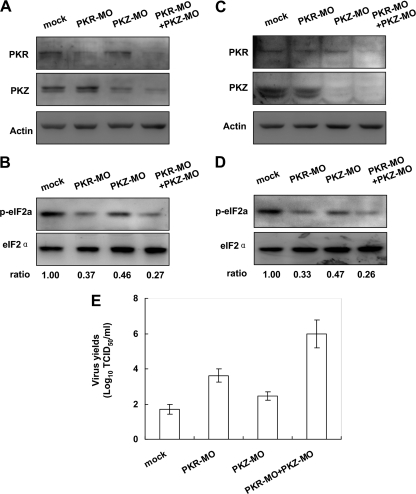
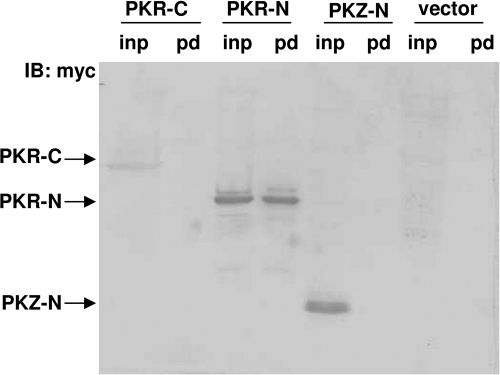
The double-stranded RNA (dsRNA)-dependent protein kinase (PKR) inhibits protein synthesis by phosphorylating eukaryotic translation initiation factor 2α (eIF2α). In fish species, in addition to PKR, there exists a PKR-like protein kinase containing Z-DNA binding domains (PKZ). However, the antiviral role of fish PKZ and the functional relationship between fish PKZ and PKR remain unknown. Here we confirmed the coexpression of fish PKZ and PKR proteins in Carassius auratus blastula embryonic (CAB) cells and identified them as two typical interferon (IFN)-inducible eIF2α kinases, both of which displayed an ability to inhibit virus replication. Strikingly, fish IFN or all kinds of IFN stimuli activated PKZ and PKR to phosphorylated eIF2α. Overexpression of both fish kinases together conferred much more significant inhibition of virus replication than overexpression of either protein, whereas morpholino knockdown of both made fish cells more vulnerable to virus infection than knockdown of either. The antiviral ability of fish PKZ was weaker than fish PKR, which correlated with its lower ability to phosphorylate eIF2α than PKR. Moreover, the independent association of fish PKZ or PKR reveals that each of them formed homodimers and that fish PKZ phosphorylated eIF2α independently on fish PKR and vice versa. These results suggest that fish PKZ and PKR play a nonredundant but cooperative role in IFN antiviral response.
Figures




Similar articles
-
Double-stranded RNA-activated protein kinase PKR of fishes and amphibians: varying the number of double-stranded RNA binding domains and lineage-specific duplications.BMC Biol. 2008 Mar 3;6:12. doi: 10.1186/1741-7007-6-12. BMC Biol. 2008. PMID: 18312693 Free PMC article.
-
The Atlantic salmon Z-DNA binding protein kinase phosphorylates translation initiation factor 2 alpha and constitutes a unique orthologue to the mammalian dsRNA-activated protein kinase R.FEBS J. 2008 Jan;275(1):184-97. doi: 10.1111/j.1742-4658.2007.06188.x. Epub 2007 Dec 11. FEBS J. 2008. PMID: 18076653
-
A PKR-like eukaryotic initiation factor 2alpha kinase from zebrafish contains Z-DNA binding domains instead of dsRNA binding domains.Proc Natl Acad Sci U S A. 2005 Feb 1;102(5):1602-7. doi: 10.1073/pnas.0408714102. Epub 2005 Jan 19. Proc Natl Acad Sci U S A. 2005. PMID: 15659550 Free PMC article.
-
PKZ, a Fish-Unique eIF2α Kinase Involved in Innate Immune Response.Front Immunol. 2020 Mar 31;11:585. doi: 10.3389/fimmu.2020.00585. eCollection 2020. Front Immunol. 2020. PMID: 32296447 Free PMC article. Review.
-
Molecular mechanisms of interferon resistance mediated by viral-directed inhibition of PKR, the interferon-induced protein kinase.Pharmacol Ther. 1998 Apr;78(1):29-46. doi: 10.1016/s0163-7258(97)00165-4. Pharmacol Ther. 1998. PMID: 9593328 Review.
Cited by
-
Adaptive duplication and genetic diversification of protein kinase R contribute to the specificity of bat-virus interactions.Sci Adv. 2022 Nov 25;8(47):eadd7540. doi: 10.1126/sciadv.add7540. Epub 2022 Nov 23. Sci Adv. 2022. PMID: 36417524 Free PMC article.
-
PKR Activation Favors Infectious Pancreatic Necrosis Virus Replication in Infected Cells.Viruses. 2016 Jun 21;8(6):173. doi: 10.3390/v8060173. Viruses. 2016. PMID: 27338445 Free PMC article.
-
Protein kinase PKR catalytic activity is required for the PKR-dependent activation of mitogen-activated protein kinases and amplification of interferon beta induction following virus infection.Virology. 2012 Jun 5;427(2):208-16. doi: 10.1016/j.virol.2012.01.029. Epub 2012 Mar 3. Virology. 2012. PMID: 22381929 Free PMC article.
-
Divergent Antiviral Mechanisms of Two Viperin Homeologs in a Recurrent Polyploid Fish.Front Immunol. 2021 Aug 31;12:702971. doi: 10.3389/fimmu.2021.702971. eCollection 2021. Front Immunol. 2021. PMID: 34531856 Free PMC article.
-
Identification of a novel Gig2 gene family specific to non-amniote vertebrates.PLoS One. 2013 Apr 4;8(4):e60588. doi: 10.1371/journal.pone.0060588. Print 2013. PLoS One. 2013. PMID: 23593256 Free PMC article.
References
-
- Al-khatib K., Williams B. R. G., Silverman R. H., Halford W., Carr D. J. J. 2003. The murine double-stranded RNA-dependent protein kinase PKR and the murine 2′,5′-oligoadenylate synthetase-dependent RNase L are required for IFN-beta-mediated resistance against herpes simplex virus type 1 in primary trigeminal ganglion culture. Virology 313:126–135 - PubMed
-
- Balachandran S., et al. 2000. Essential role for the dsRNA-dependent protein kinase PKR in innate immunity to viral infection. Immunity 13:129–141 - PubMed
-
- Bergan V., Jagus R., Lauksund S., Kileng Ø., Robertsen B. 2008. The Atlantic salmon Z-DNA binding protein kinase phosphorylates translation initiation factor 2 alpha and constitutes a unique orthologue to the mammalian dsRNA-activated protein kinase R. FEBS J. 275:184–197 - PubMed
-
- Cai R., Williams B. R. G. 1998. Mutations in the double-stranded RNA-activated protein kinase insert region that uncouple catalysis from eIF2α binding. J. Biol. Chem. 273:11274–11280 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
