

At a crossroads: human DNA tumor viruses and the host DNA damage response
- PMID: 21927617
- PMCID: PMC3171950
- DOI: 10.2217/fvl.11.55
At a crossroads: human DNA tumor viruses and the host DNA damage response
Abstract
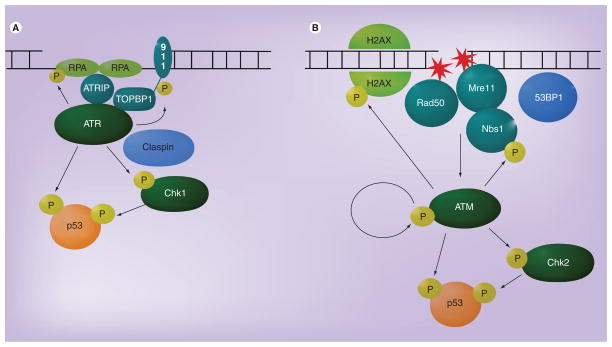
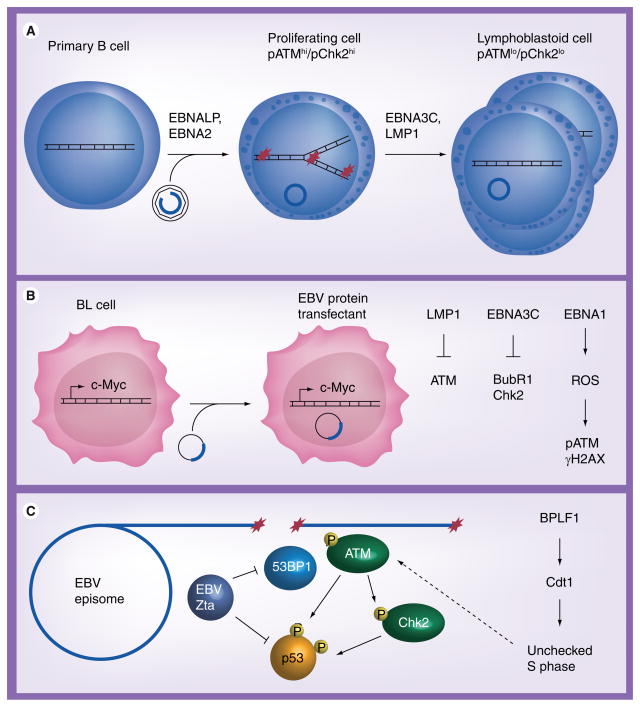
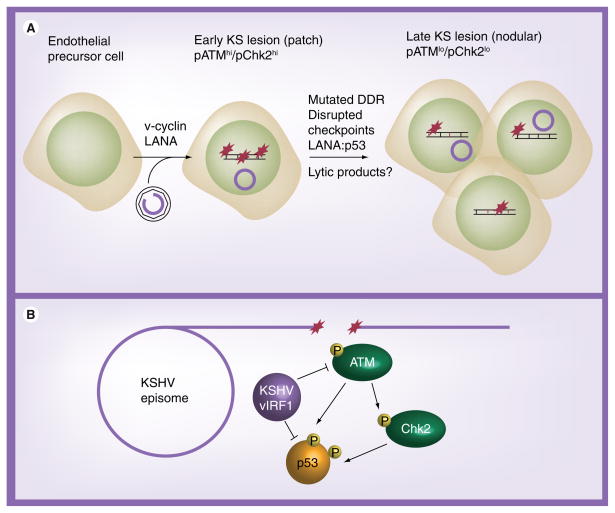
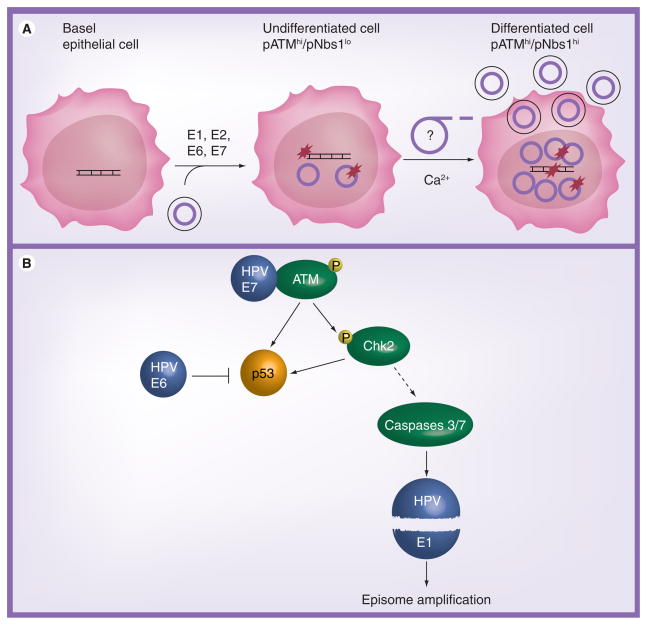
Human DNA tumor viruses induce host cell proliferation in order to establish the necessary cellular milieu to replicate viral DNA. The consequence of such viral-programmed induction of proliferation coupled with the introduction of foreign replicating DNA structures makes these viruses particularly sensitive to the host DNA damage response machinery. In fact, sensors of DNA damage are often activated and modulated by DNA tumor viruses in both latent and lytic infection. This article focuses on the role of the DNA damage response during the life cycle of human DNA tumor viruses, with a particular emphasis on recent advances in our understanding of the role of the DNA damage response in EBV, Kaposi's sarcoma-associated herpesvirus and human papillomavirus infection.
Figures
Similar articles
-
Interplay between DNA tumor viruses and the host DNA damage response.Curr Top Microbiol Immunol. 2013;371:229-57. doi: 10.1007/978-3-642-37765-5_9. Curr Top Microbiol Immunol. 2013. PMID: 23686238 Free PMC article. Review.
-
Kaposi's Sarcoma-Associated Herpesvirus Lytic Replication Is Independent of Anaphase-Promoting Complex Activity.J Virol. 2020 Jun 16;94(13):e02079-19. doi: 10.1128/JVI.02079-19. Print 2020 Jun 16. J Virol. 2020. PMID: 32295923 Free PMC article.
-
Interplay Between KSHV and the Host DNA Damage Response.Front Cell Infect Microbiol. 2020 Dec 9;10:604351. doi: 10.3389/fcimb.2020.604351. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33425783 Free PMC article. Review.
-
Complex Interactions between Cohesin and CTCF in Regulation of Kaposi's Sarcoma-Associated Herpesvirus Lytic Transcription.J Virol. 2020 Jan 6;94(2):e01279-19. doi: 10.1128/JVI.01279-19. Print 2020 Jan 6. J Virol. 2020. PMID: 31666380 Free PMC article.
-
Full-Length Isoforms of Kaposi's Sarcoma-Associated Herpesvirus Latency-Associated Nuclear Antigen Accumulate in the Cytoplasm of Cells Undergoing the Lytic Cycle of Replication.J Virol. 2017 Nov 30;91(24):e01532-17. doi: 10.1128/JVI.01532-17. Print 2017 Dec 15. J Virol. 2017. PMID: 28978712 Free PMC article.
Cited by
-
SARS-CoV-2 and the DNA damage response.J Gen Virol. 2023 Nov;104(11):001918. doi: 10.1099/jgv.0.001918. J Gen Virol. 2023. PMID: 37948194 Free PMC article. Review.
-
Histone deacetylase classes I and II regulate Kaposi's sarcoma-associated herpesvirus reactivation.J Virol. 2014 Jan;88(2):1281-92. doi: 10.1128/JVI.02665-13. Epub 2013 Nov 13. J Virol. 2014. PMID: 24227836 Free PMC article.
-
Viral reprogramming of host transcription initiation.Nucleic Acids Res. 2024 May 22;52(9):5016-5032. doi: 10.1093/nar/gkae175. Nucleic Acids Res. 2024. PMID: 38471819 Free PMC article.
-
Interplay between DNA tumor viruses and the host DNA damage response.Curr Top Microbiol Immunol. 2013;371:229-57. doi: 10.1007/978-3-642-37765-5_9. Curr Top Microbiol Immunol. 2013. PMID: 23686238 Free PMC article. Review.
-
Marek's Disease Virus Disables the ATR-Chk1 Pathway by Activating STAT3.J Virol. 2019 Apr 17;93(9):e02290-18. doi: 10.1128/JVI.02290-18. Print 2019 May 1. J Virol. 2019. PMID: 30787154 Free PMC article.
References
-
- Bartkova J, Rezaei N, Liontos M, et al. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature. 2006;444(7119):633–637. Along with [2], demonstrates that the functional outcome of oncogene-induced DNA damage response (DDR) activation is senescence. These data support the model whereby ataxia-telangiectasia mutated (ATM)/ataxia-telangiectasia RAD3-related (ATR) activation downstream of replicative stress acts as an early barrier to tumorigenesis. - PubMed
-
- Di Micco R, Fumagalli M, Cicalese A, et al. Oncogene-induced senescence is a DNA damage response triggered by DNA hyper-replication. Nature. 2006;444(7119):638–642. Along with [1], demonstrates that the functional outcome of oncogene-induced DDR activation is senescence. These data support the model whereby ATM/ATR activation downstream of replicative stress acts as an early barrier to tumorigenesis. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434(7035):864–870. - PubMed
-
- Gorgoulis VG, Vassiliou LV, Karakaidos P, et al. Activation of the DNA damage checkpoint and genomic instability in human precancerous lesions. Nature. 2005;434(7035):907–913. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources