

Dysregulation of astrocyte-motoneuron cross-talk in mutant superoxide dismutase 1-related amyotrophic lateral sclerosis
- PMID: 21908873
- PMCID: PMC3170534
- DOI: 10.1093/brain/awr193
Dysregulation of astrocyte-motoneuron cross-talk in mutant superoxide dismutase 1-related amyotrophic lateral sclerosis
Abstract
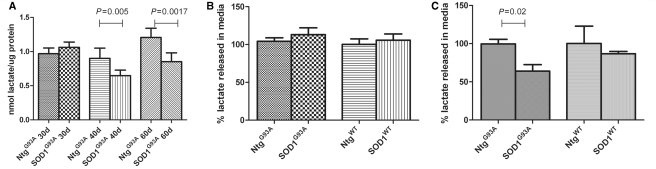
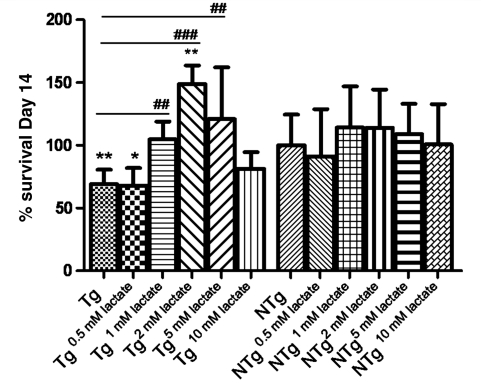
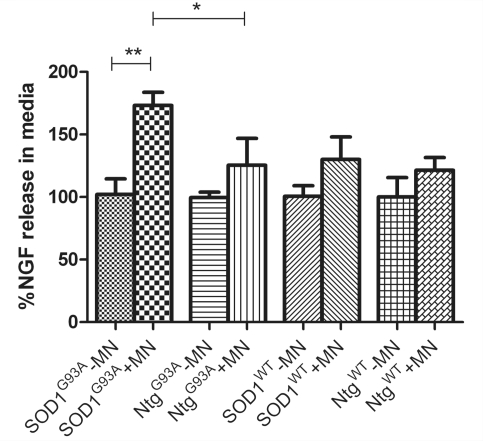
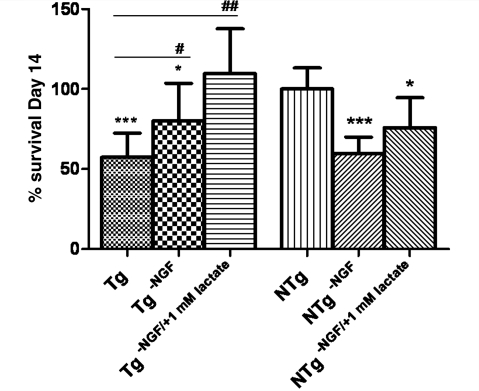
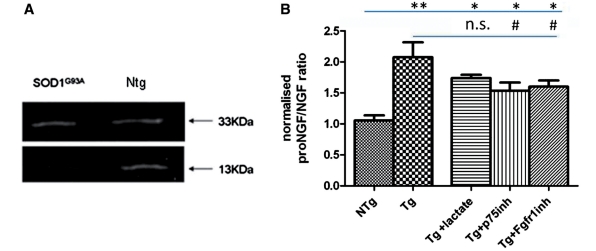
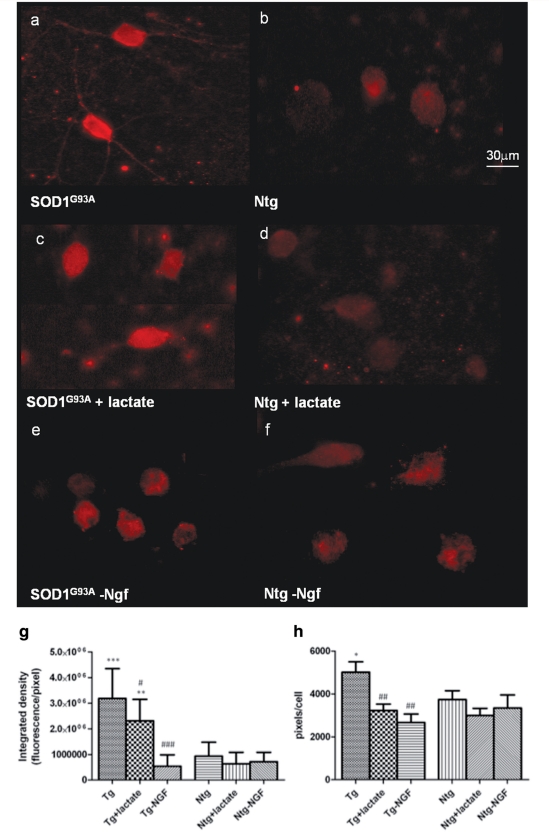
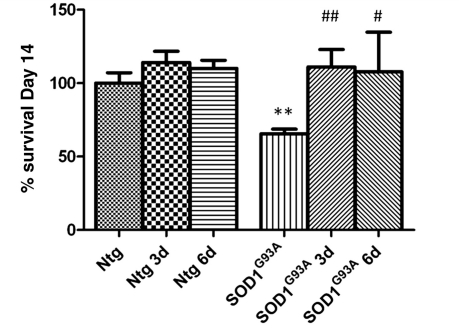
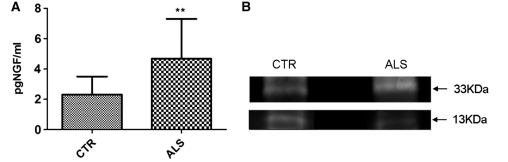
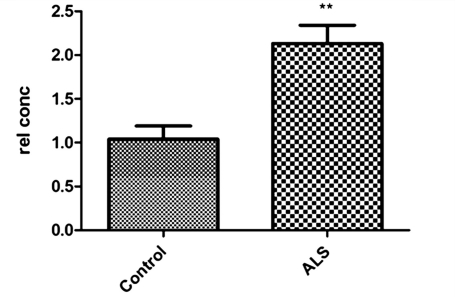
Amyotrophic lateral sclerosis is a neurodegenerative disease in which death of motoneurons leads to progressive failure of the neuromuscular system resulting in death frequently within 2-3 years of symptom onset. Focal onset and propagation of the disease symptoms to contiguous motoneuron groups is a striking feature of the human disease progression. Recent work, using mutant superoxide dismutase 1 murine models and in vitro culture systems has indicated that astrocytes are likely to contribute to the propagation of motoneuron injury and disease progression. However, the basis of this astrocyte toxicity and/or failure of motoneuron support has remained uncertain. Using a combination of in vivo and in vitro model systems of superoxide dismutase 1-related amyotrophic lateral sclerosis, linked back to human biosamples, we set out to elucidate how astrocyte properties change in the presence of mutant superoxide dismutase 1 to contribute to motoneuron injury. Gene expression profiling of spinal cord astrocytes from presymptomatic transgenic mice expressing mutant superoxide dismutase 1 revealed two striking changes. First, there was evidence of metabolic dysregulation and, in particular, impairment of the astrocyte lactate efflux transporter, with resultant decrease of spinal cord lactate levels. Second, there was evidence of increased nerve growth factor production and dysregulation of the ratio of pro-nerve growth factor to mature nerve growth factor, favouring p75 receptor expression and activation by neighbouring motoneurons. Functional in vitro studies showed that astrocytes expressing mutant superoxide dismutase 1 are toxic to normal motoneurons. We provide evidence that reduced metabolic support from lactate release and activation of pro-nerve growth factor-p75 receptor signalling are key components of this toxicity. Preservation of motoneuron viability could be achieved by increasing lactate provision to motoneurons, depletion of increased pro-nerve growth factor levels or p75 receptor blockade. These findings are likely to be relevant to human amyotrophic lateral sclerosis, where we have demonstrated increased levels of pro-nerve growth factor in cerebrospinal fluid and increased expression of the p75 receptor by spinal motoneurons. Taken together, these data confirm that altered properties of astrocytes are likely to play a crucial role in the propagation of motoneuron injury in superoxide dismutase 1-related amyotrophic lateral sclerosis and indicate that manipulation of the energy supply to motoneurons as well as inhibition of p75 receptor signalling may represent valuable neuroprotective strategies.
Figures
Similar articles
-
Expression of the low affinity neurotrophin receptor p75 in spinal motoneurons in a transgenic mouse model for amyotrophic lateral sclerosis.Neuroscience. 2003;116(3):685-94. doi: 10.1016/s0306-4522(02)00755-8. Neuroscience. 2003. PMID: 12573711
-
Extracellular ATP and the P2X7 receptor in astrocyte-mediated motor neuron death: implications for amyotrophic lateral sclerosis.J Neuroinflammation. 2010 Jun 9;7:33. doi: 10.1186/1742-2094-7-33. J Neuroinflammation. 2010. PMID: 20534165 Free PMC article.
-
Astrocytic production of nerve growth factor in motor neuron apoptosis: implications for amyotrophic lateral sclerosis.J Neurochem. 2004 Apr;89(2):464-73. doi: 10.1111/j.1471-4159.2004.02357.x. J Neurochem. 2004. PMID: 15056289
-
Complexity of astrocyte-motor neuron interactions in amyotrophic lateral sclerosis.Neurodegener Dis. 2005;2(3-4):139-46. doi: 10.1159/000089619. Neurodegener Dis. 2005. PMID: 16909019 Review.
-
Inhibitory synaptic regulation of motoneurons: a new target of disease mechanisms in amyotrophic lateral sclerosis.Mol Neurobiol. 2012 Feb;45(1):30-42. doi: 10.1007/s12035-011-8217-x. Epub 2011 Nov 10. Mol Neurobiol. 2012. PMID: 22072396 Free PMC article. Review.
Cited by
-
P75 and phosphorylated c-Jun are differentially regulated in spinal motoneurons following axotomy in rats.Neural Regen Res. 2012 Sep 15;7(26):2005-11. doi: 10.3969/j.issn.1673-5374.2012.26.001. Neural Regen Res. 2012. PMID: 25624831 Free PMC article.
-
Transcriptome Profiling Following Neuronal and Glial Expression of ALS-Linked SOD1 in Drosophila.G3 (Bethesda). 2013 Apr 9;3(4):695-708. doi: 10.1534/g3.113.005850. G3 (Bethesda). 2013. PMID: 23550139 Free PMC article.
-
Astrocytes drive upregulation of the multidrug resistance transporter ABCB1 (P-Glycoprotein) in endothelial cells of the blood-brain barrier in mutant superoxide dismutase 1-linked amyotrophic lateral sclerosis.Glia. 2016 Aug;64(8):1298-313. doi: 10.1002/glia.23003. Epub 2016 May 9. Glia. 2016. PMID: 27158936 Free PMC article.
-
Astrocytes in neurodegenerative disease.Cold Spring Harb Perspect Biol. 2015 Apr 15;7(6):a020628. doi: 10.1101/cshperspect.a020628. Cold Spring Harb Perspect Biol. 2015. PMID: 25877220 Free PMC article. Review.
-
Regional Differences in Heat Shock Protein 25 Expression in Brain and Spinal Cord Astrocytes of Wild-Type and SOD1 G93A Mice.Cells. 2021 May 19;10(5):1257. doi: 10.3390/cells10051257. Cells. 2021. PMID: 34069691 Free PMC article.
References
-
- Bierl MA, Jones EE, Crutcher KA, Isaacson LG. ‘Mature’ nerve growth factor is a minor species in most peripheral tissues. Neurosci Lett. 2005;380:133–7. - PubMed
-
- Bilsland LG, Nirmalananthan N, Yip J, Greensmith L, Duchen MR. Expression of mutant SOD1 in astrocytes induces functional deficits in motoneuron mitochondria. J Neurochem. 2008;107:1271–83. - PubMed
-
- Boillee S, Yamanaka K, Lobsiger CS, Copeland NG, Jenkins NA, Kassiotis G, et al. Onset and progression in inherited ALS determined by motor neurons and microglia. Science. 2006;312:1389–92. - PubMed
-
- Boyd JG, Gordon T. The neurotrophin receptors, trkB and p75, differentially regulate motor axonal regeneration. J Neurobiol. 2001;49:314–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
