

Absence of cross-presenting cells in the salivary gland and viral immune evasion confine cytomegalovirus immune control to effector CD4 T cells
- PMID: 21901102
- PMCID: PMC3161985
- DOI: 10.1371/journal.ppat.1002214
Absence of cross-presenting cells in the salivary gland and viral immune evasion confine cytomegalovirus immune control to effector CD4 T cells
Abstract
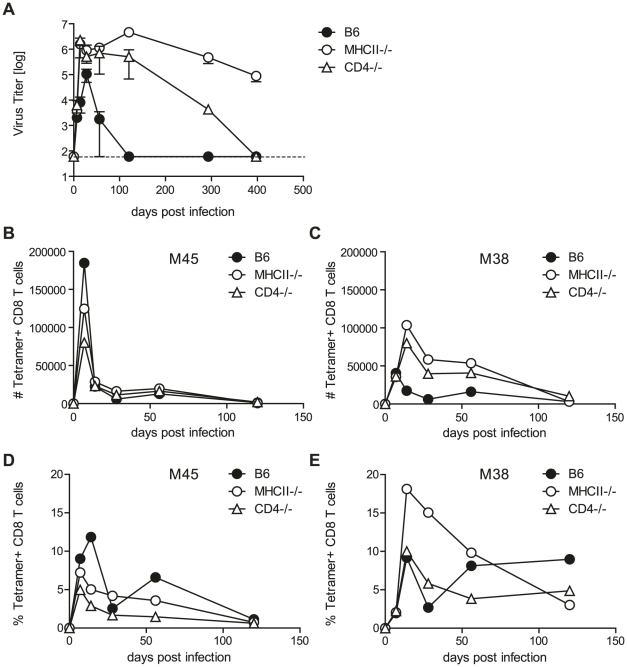
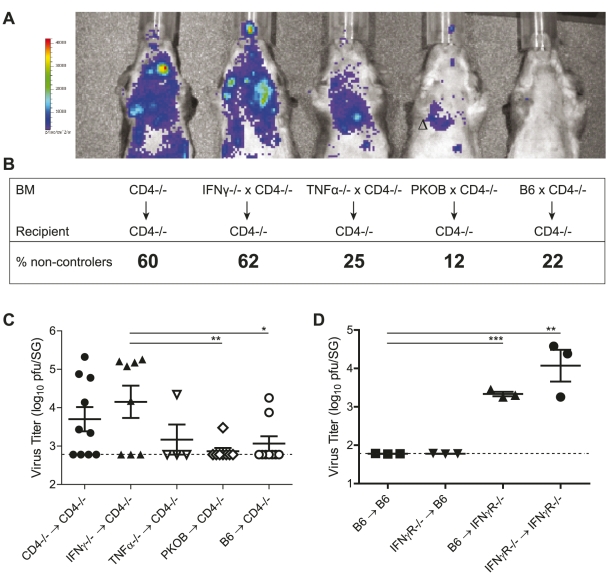
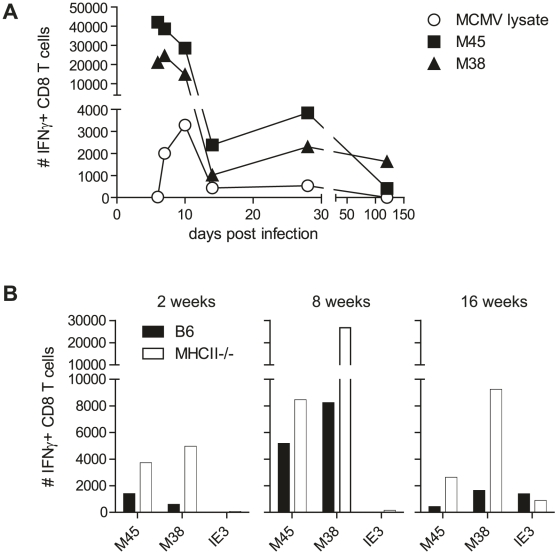
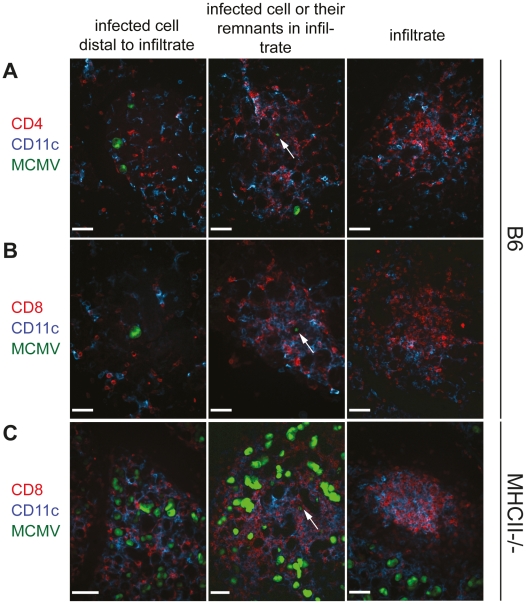
Horizontal transmission of cytomegaloviruses (CMV) occurs via prolonged excretion from mucosal surfaces. We used murine CMV (MCMV) infection to investigate the mechanisms of immune control in secretory organs. CD4 T cells were crucial to cease MCMV replication in the salivary gland (SG) via direct secretion of IFNγ that initiated antiviral signaling on non-hematopoietic cells. In contrast, CD4 T cell helper functions for CD8 T cells or B cells were dispensable. Despite SG-resident MCMV-specific CD8 T cells being able to produce IFNγ, the absence of MHC class I molecules on infected acinar glandular epithelial cells due to viral immune evasion, and the paucity of cross-presenting antigen presenting cells (APCs) prevented their local activation. Thus, local activation of MCMV-specific T cells is confined to the CD4 subset due to exclusive presentation of MCMV-derived antigens by MHC class II molecules on bystander APCs, resulting in IFNγ secretion interfering with viral replication in cells of non-hematopoietic origin.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Hematopoietic cell-mediated dissemination of murine cytomegalovirus is regulated by NK cells and immune evasion.PLoS Pathog. 2021 Jan 28;17(1):e1009255. doi: 10.1371/journal.ppat.1009255. eCollection 2021 Jan. PLoS Pathog. 2021. PMID: 33508041 Free PMC article.
-
Salivary gland resident APCs are Flt3L- and CCR2-independent macrophage-like cells incapable of cross-presentation.Eur J Immunol. 2014 Mar;44(3):706-14. doi: 10.1002/eji.201343992. Epub 2013 Dec 23. Eur J Immunol. 2014. PMID: 24271944
-
Priming of CD8+ T cells against cytomegalovirus-encoded antigens is dominated by cross-presentation.J Immunol. 2013 Mar 15;190(6):2767-77. doi: 10.4049/jimmunol.1200966. Epub 2013 Feb 6. J Immunol. 2013. PMID: 23390296
-
MHC class I immune evasion in MCMV infection.Med Microbiol Immunol. 2008 Jun;197(2):191-204. doi: 10.1007/s00430-008-0089-y. Epub 2008 Mar 11. Med Microbiol Immunol. 2008. PMID: 18330598 Review.
-
The salivary glands as a privileged site of cytomegalovirus immune evasion and persistence.Med Microbiol Immunol. 2008 Jun;197(2):205-13. doi: 10.1007/s00430-008-0077-2. Epub 2008 Feb 8. Med Microbiol Immunol. 2008. PMID: 18259775 Review.
Cited by
-
Control of murine cytomegalovirus infection by γδ T cells.PLoS Pathog. 2015 Feb 6;11(2):e1004481. doi: 10.1371/journal.ppat.1004481. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25658831 Free PMC article.
-
Spatial kinetics and immune control of murine cytomegalovirus infection in the salivary glands.PLoS Comput Biol. 2024 Aug 16;20(8):e1011940. doi: 10.1371/journal.pcbi.1011940. eCollection 2024 Aug. PLoS Comput Biol. 2024. PMID: 39150988 Free PMC article.
-
Molecular investigation of the 7.2 kb RNA of murine cytomegalovirus.Virol J. 2013 Dec 2;10:348. doi: 10.1186/1743-422X-10-348. Virol J. 2013. PMID: 24295514 Free PMC article.
-
Hematopoietic cell-mediated dissemination of murine cytomegalovirus is regulated by NK cells and immune evasion.PLoS Pathog. 2021 Jan 28;17(1):e1009255. doi: 10.1371/journal.ppat.1009255. eCollection 2021 Jan. PLoS Pathog. 2021. PMID: 33508041 Free PMC article.
-
CD4+ but not CD8+ T cells are required for protection against severe guinea pig cytomegalovirus infections.PLoS Pathog. 2024 Nov 4;20(11):e1012515. doi: 10.1371/journal.ppat.1012515. eCollection 2024 Nov. PLoS Pathog. 2024. PMID: 39495799 Free PMC article.
References
-
- Mocarski ES, Shenk T, Pass RF. Cytomegaloviruses; In: Howley DKKaPM., editor. Philadelphia: Lippincott, Williams & Wilkins; 2007.
-
- Boppana SB, Fowler KB. Persistence in the population: epidemiology and transmisson. In: Arvin A C-FG, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K, editors. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis. Cambridge: Cambridge University Press; 2007. 2011/02/25 ed.
-
- Britt W. Virus entry into host, establishment of infection, spread in host, mechanisms of tissue damage. In: Arvin A C-FG, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K, editors. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis. Cambridge: Cambridge University Press; 2007. 2011/02/25 ed. - PubMed
-
- Campbell AE, Cavanaugh VJ, Slater JS. The salivary glands as a privileged site of cytomegalovirus immune evasion and persistence. Med Microbiol Immunol. 2008;197:205–213. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
