

Individual axons regulate the myelinating potential of single oligodendrocytes in vivo
- PMID: 21880787
- PMCID: PMC3177314
- DOI: 10.1242/dev.071001
Individual axons regulate the myelinating potential of single oligodendrocytes in vivo
Abstract
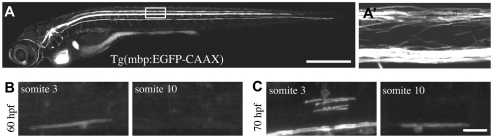
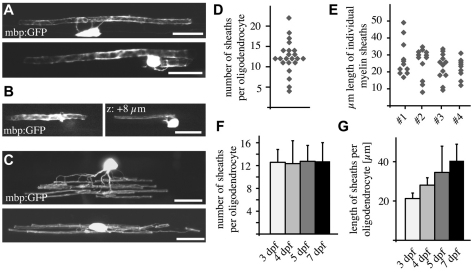
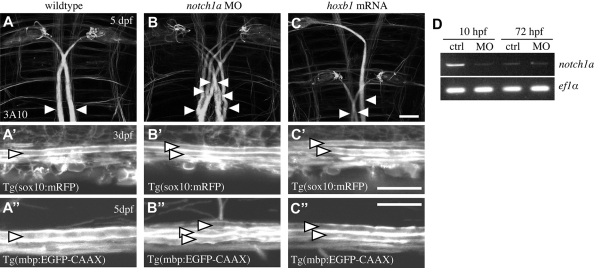
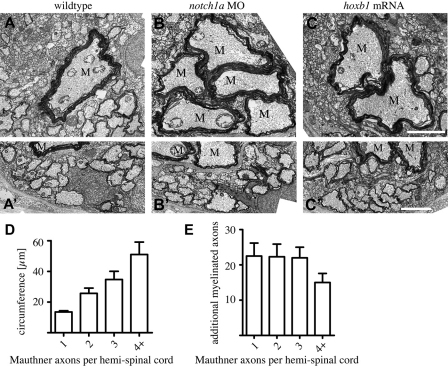
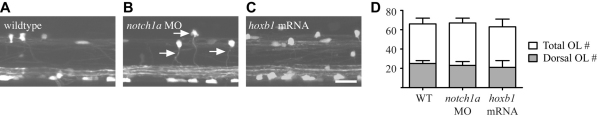
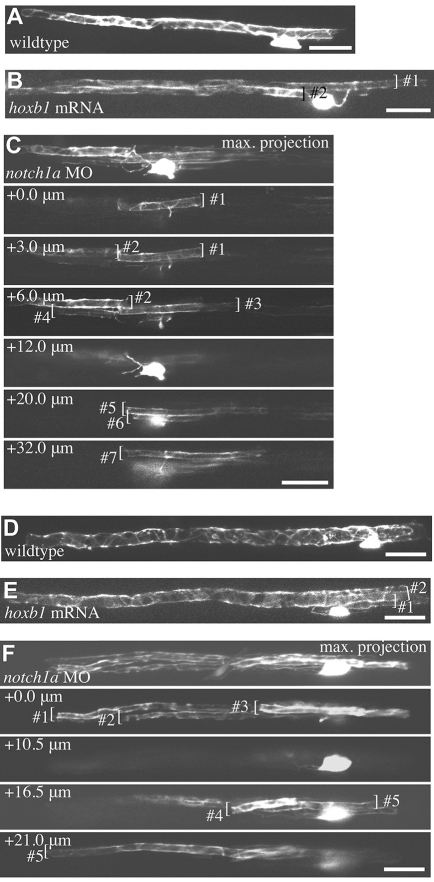
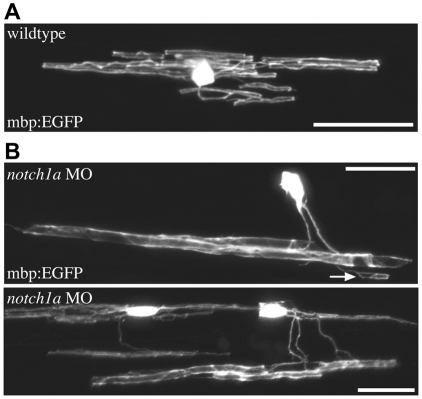
The majority of axons in the central nervous system (CNS) are eventually myelinated by oligodendrocytes, but whether the timing and extent of myelination in vivo reflect intrinsic properties of oligodendrocytes, or are regulated by axons, remains undetermined. Here, we use zebrafish to study CNS myelination at single-cell resolution in vivo. We show that the large caliber Mauthner axon is the first to be myelinated (shortly before axons of smaller caliber) and that the presence of supernumerary large caliber Mauthner axons can profoundly affect myelination by single oligodendrocytes. Oligodendrocytes that typically myelinate just one Mauthner axon in wild type can myelinate multiple supernumerary Mauthner axons. Furthermore, oligodendrocytes that exclusively myelinate numerous smaller caliber axons in wild type can readily myelinate small caliber axons in addition to the much larger caliber supernumerary Mauthner axons. These data indicate that single oligodendrocytes can myelinate diverse axons and that their myelinating potential is actively regulated by individual axons.
Figures
Similar articles
-
Myelination of Neuronal Cell Bodies when Myelin Supply Exceeds Axonal Demand.Curr Biol. 2018 Apr 23;28(8):1296-1305.e5. doi: 10.1016/j.cub.2018.02.068. Epub 2018 Apr 5. Curr Biol. 2018. PMID: 29628374 Free PMC article.
-
Oligodendrocytes are not inherently programmed to myelinate a specific size of axon.J Comp Neurol. 1998 Sep 14;399(1):94-100. J Comp Neurol. 1998. PMID: 9725703
-
Oligodendrocyte Development in the Absence of Their Target Axons In Vivo.PLoS One. 2016 Oct 7;11(10):e0164432. doi: 10.1371/journal.pone.0164432. eCollection 2016. PLoS One. 2016. PMID: 27716830 Free PMC article.
-
Axon myelination. Myelination without myelin-associated glycoprotein.Curr Biol. 1994 Sep 1;4(9):847-50. doi: 10.1016/s0960-9822(00)00190-1. Curr Biol. 1994. PMID: 7529638 Review.
-
Insights into mechanisms of central nervous system myelination using zebrafish.Glia. 2016 Mar;64(3):333-49. doi: 10.1002/glia.22897. Epub 2015 Aug 6. Glia. 2016. PMID: 26250418 Review.
Cited by
-
Olig1 function is required for oligodendrocyte differentiation in the mouse brain.J Neurosci. 2015 Mar 11;35(10):4386-402. doi: 10.1523/JNEUROSCI.4962-14.2015. J Neurosci. 2015. PMID: 25762682 Free PMC article.
-
Developmental Changes in Oligodendrocyte Genesis, Myelination, and Associated Behavioral Dysfunction in a Rat Model of Intra-generational Protein Malnutrition.Mol Neurobiol. 2019 Jan;56(1):595-610. doi: 10.1007/s12035-018-1065-1. Epub 2018 May 12. Mol Neurobiol. 2019. PMID: 29752656
-
Imaging Myelination In Vivo Using Transparent Animal Models.Brain Plast. 2016 Dec 21;2(1):3-29. doi: 10.3233/BPL-160029. Brain Plast. 2016. PMID: 29765846 Free PMC article. Review.
-
Thyroid hormone deficiency during zebrafish development impairs central nervous system myelination.PLoS One. 2021 Aug 17;16(8):e0256207. doi: 10.1371/journal.pone.0256207. eCollection 2021. PLoS One. 2021. PMID: 34403440 Free PMC article.
-
Spinal cord precursors utilize neural crest cell mechanisms to generate hybrid peripheral myelinating glia.Elife. 2021 Feb 8;10:e64267. doi: 10.7554/eLife.64267. Elife. 2021. PMID: 33554855 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
