

Novel C-terminal motif within Sec7 domain of guanine nucleotide exchange factors regulates ADP-ribosylation factor (ARF) binding and activation
- PMID: 21828055
- PMCID: PMC3196086
- DOI: 10.1074/jbc.M111.230631
Novel C-terminal motif within Sec7 domain of guanine nucleotide exchange factors regulates ADP-ribosylation factor (ARF) binding and activation
Abstract
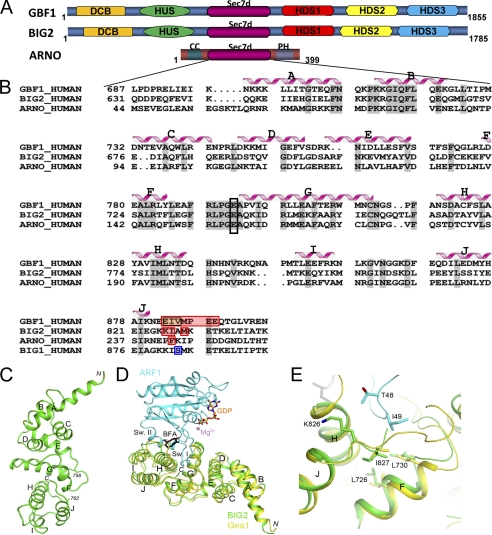
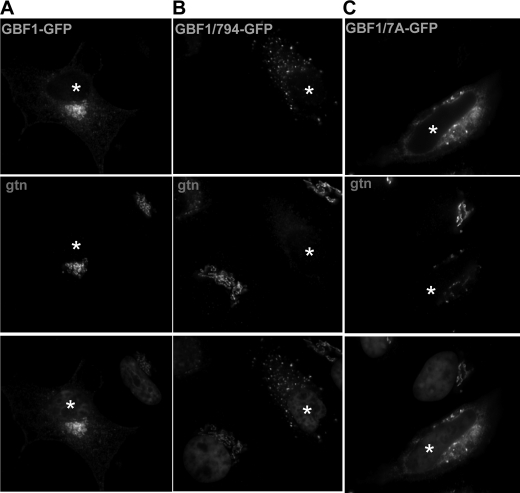
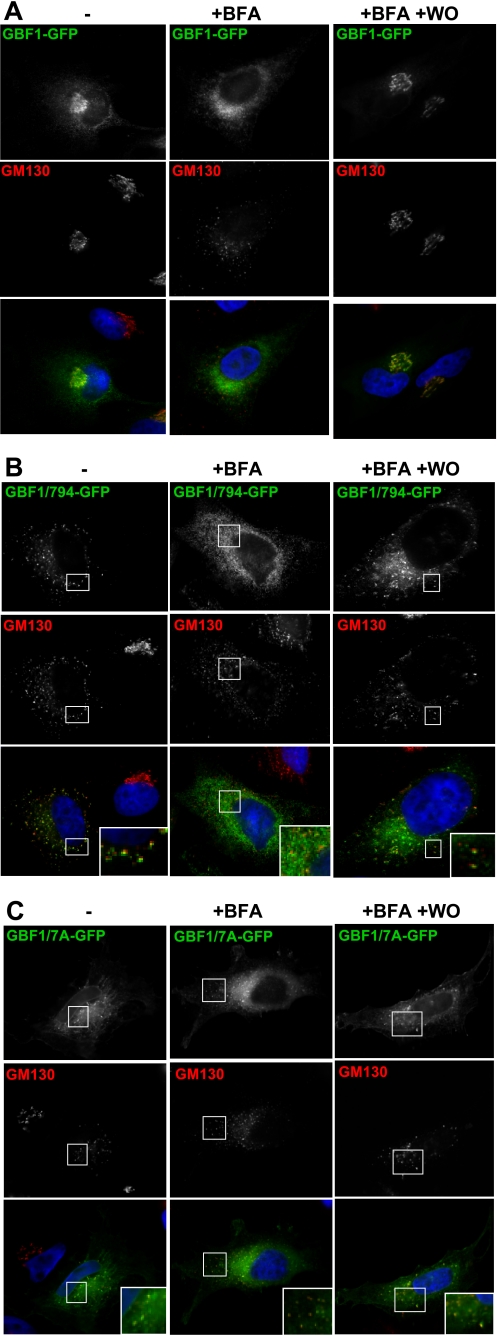
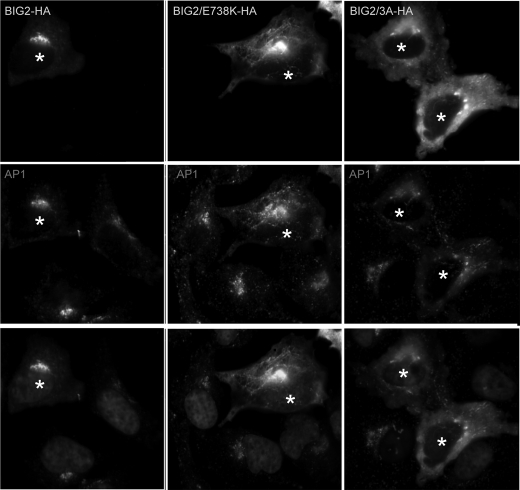
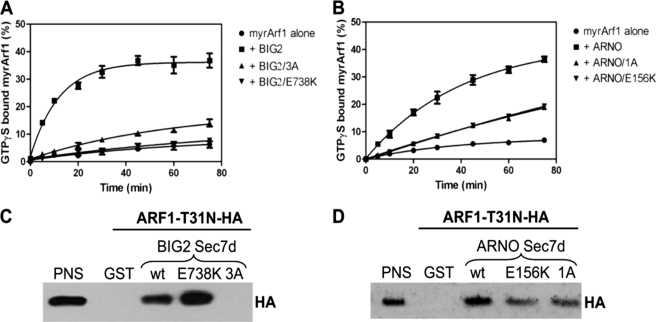
ADP-ribosylation factors (ARFs) and their activating guanine nucleotide exchange factors (GEFs) play key roles in membrane traffic and signaling. All ARF GEFs share a ∼200-residue Sec7 domain (Sec7d) that alone catalyzes the GDP to GTP exchange that activates ARF. We determined the crystal structure of human BIG2 Sec7d. A C-terminal loop immediately following helix J (loop>J) was predicted to form contacts with helix H and the switch I region of the cognate ARF, suggesting that loop>J may participate in the catalytic reaction. Indeed, we identified multiple alanine substitutions within loop>J of the full length and/or Sec7d of two large brefeldin A-sensitive GEFs (GBF1 and BIG2) and one small brefeldin A-resistant GEF (ARNO) that abrogated binding of ARF and a single alanine substitution that allowed ARF binding but inhibited GDP to GTP exchange. Loop>J sequences are highly conserved, suggesting that loop>J plays a crucial role in the catalytic activity of all ARF GEFs. Using GEF mutants unable to bind ARF, we showed that GEFs associate with membranes independently of ARF and catalyze ARF activation in vivo only when membrane-associated. Our structural, cell biological, and biochemical findings identify loop>J as a key regulatory motif essential for ARF binding and GDP to GTP exchange by GEFs and provide evidence for the requirement of membrane association during GEF activity.
Figures
Similar articles
-
Promiscuity of the catalytic Sec7 domain within the guanine nucleotide exchange factor GBF1 in ARF activation, Golgi homeostasis, and effector recruitment.Mol Biol Cell. 2019 Jun 1;30(12):1523-1535. doi: 10.1091/mbc.E18-11-0711. Epub 2019 Apr 3. Mol Biol Cell. 2019. PMID: 30943106 Free PMC article.
-
C-terminal motif within Sec7 domain regulates guanine nucleotide exchange activity via tuning protein conformation.Biochem Biophys Res Commun. 2014 Mar 28;446(1):380-6. doi: 10.1016/j.bbrc.2014.02.125. Epub 2014 Mar 5. Biochem Biophys Res Commun. 2014. PMID: 24613384
-
EFA6 controls Arf1 and Arf6 activation through a negative feedback loop.Proc Natl Acad Sci U S A. 2014 Aug 26;111(34):12378-83. doi: 10.1073/pnas.1409832111. Epub 2014 Aug 11. Proc Natl Acad Sci U S A. 2014. PMID: 25114232 Free PMC article.
-
Allosteric regulation of Arf GTPases and their GEFs at the membrane interface.Small GTPases. 2016 Oct;7(4):283-296. doi: 10.1080/21541248.2016.1215778. Epub 2016 Jul 22. Small GTPases. 2016. PMID: 27449855 Free PMC article. Review.
-
Arf, Sec7 and Brefeldin A: a model towards the therapeutic inhibition of guanine nucleotide-exchange factors.Biochem Soc Trans. 2005 Dec;33(Pt 6):1265-8. doi: 10.1042/BST0331265. Biochem Soc Trans. 2005. PMID: 16246094 Review.
Cited by
-
The pleckstrin homology (PH) domain of the Arf exchange factor Brag2 is an allosteric binding site.J Biol Chem. 2012 Jul 13;287(29):24273-83. doi: 10.1074/jbc.M112.368084. Epub 2012 May 21. J Biol Chem. 2012. PMID: 22613714 Free PMC article.
-
The cytohesin paralog Sec7 of Dictyostelium discoideum is required for phagocytosis and cell motility.Cell Commun Signal. 2013 Aug 1;11:54. doi: 10.1186/1478-811X-11-54. Cell Commun Signal. 2013. PMID: 23915312 Free PMC article.
-
The Guanine Nucleotide Exchange Factor GBF1 Participates in Rotavirus Replication.J Virol. 2019 Sep 12;93(19):e01062-19. doi: 10.1128/JVI.01062-19. Print 2019 Oct 1. J Virol. 2019. PMID: 31270230 Free PMC article.
-
Highly conserved motifs within the large Sec7 ARF guanine nucleotide exchange factor GBF1 target it to the Golgi and are critical for GBF1 activity.Am J Physiol Cell Physiol. 2018 Jun 1;314(6):C675-C689. doi: 10.1152/ajpcell.00221.2017. Epub 2018 Feb 14. Am J Physiol Cell Physiol. 2018. PMID: 29443553 Free PMC article.
-
Coordinated Activation of ARF1 GTPases by ARF-GEF GNOM Dimers Is Essential for Vesicle Trafficking in Arabidopsis.Plant Cell. 2020 Aug;32(8):2491-2507. doi: 10.1105/tpc.20.00240. Epub 2020 Jun 2. Plant Cell. 2020. PMID: 32487565 Free PMC article.
References
-
- Jackson C. L., Casanova J. E. (2000) Trends Cell Biol. 10, 60–67 - PubMed
-
- Chardin P., Paris S., Antonny B., Robineau S., Béraud-Dufour S., Jackson C. L., Chabre M. (1996) Nature 384, 481–484 - PubMed
-
- Antonny B., Beraud-Dufour S., Chardin P., Chabre M. (1997) Biochemistry 36, 4675–4684 - PubMed
-
- Mossessova E., Gulbis J. M., Goldberg J. (1998) Cell 92, 415–423 - PubMed
-
- Cherfils J., Ménétrey J., Mathieu M., Le Bras G., Robineau S., Béraud-Dufour S., Antonny B., Chardin P. (1998) Nature 392, 101–105 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
