

Review
doi: 10.1038/nrmicro2613.
Influenza A viruses: new research developments
Affiliations
- PMID: 21747392
- PMCID: PMC10433403
- DOI: 10.1038/nrmicro2613
Item in Clipboard
Review
Influenza A viruses: new research developments
Nat Rev Microbiol.
.
Abstract
Influenza A viruses are zoonotic pathogens that continuously circulate and change in several animal hosts, including birds, pigs, horses and humans. The emergence of novel virus strains that are capable of causing human epidemics or pandemics is a serious possibility. Here, we discuss the value of surveillance and characterization of naturally occurring influenza viruses, and review the impact that new developments in the laboratory have had on our understanding of the host tropism and virulence of viruses. We also revise the lessons that have been learnt from the pandemic viruses of the past 100 years.
Figures

a ∣ The influenza A virus genome consists of eight single-stranded RNAs that encode 11 or 12 proteins. These are nuclear export protein (NEP; also known as NS2) and the host antiviral response antagonist non-structural protein 1 (NS1), which are encoded by the NS segment; the matrix protein M1 and the ion channel M2, which are encoded by the M segment; the receptor-binding protein haemagglutinin (HA), the sialic acid-destroying enzyme neuraminidase (NA), nucleoprotein (NP), and the components of the RNA-dependent RNA polymerase complex (PB1, PB2 and PA), all expressed from their respective genome segments; and the newly identified N40 protein, which is expressed from the PB1 segment and has an unknown function. In addition, some viruses express the pro-apoptotic protein PB1-F2, which is encoded by a second ORF in the PB1 segment. Within the virion, each of the eight viral segments forms a viral ribonucleoprotein (RNP) complex: viral RNA is wrapped around NP, and this structure is then bound to the viral polymerase complex. b ∣ The antigenic properties of HA allow the classification of influenza A viruses into two major groups, 1 and 2, which are further classified into five clades and 16 subtypes. c ∣ In the initial stages of influenza A virus replication, the viral HA attaches to host cell receptors that contain terminal α-2,6-linked or α-2-3-linked sialic acid (α-2,6-SA or α-2,3-SA) moieties, and the virus enters the cell by receptor-mediated endocytosis. Cleavage of HA by cellular proteases is required to expose the HA peptide that is responsible for the fusion between the viral envelope and the endosomal membrane (see below). Acidification of the endocytic vesicle opens the M2 ion channel, resulting in acidification of the inside of the virion, a process that is required for proper uncoating of the RNP complexes that contain the viral genome. Acidification of the endosome also triggers the pH-dependent fusion step that is mediated by HA and results in the cytoplasmic release of the RNP complexes. These translocate to the nucleus, where the RNA-dependent RNA polymerase transcribes and replicates the negative-sense viral RNA ((−)vRNA), giving rise to three types of RNA molecules: the complementary positive-sense RNA ((+)cRNA), which it uses as a template to generate more vRNA; negative-sense small viral RNAs (svRNAs), which are thought to regulate the switch from transcription to replication,; and the viral mRNAs, which are exported to the cytoplasm for translation. Viral proteins that are needed in replication and transcription are translocated back to the nucleus, and progeny RNPs are then exported to the cytoplasm for packaging, assisted by M1 and NEP. Viral HA, NA and M2 are transported by the trans-Golgi secretory pathway, and the mature proteins arrive at the plasma membrane, where M1 assists in the formation of virus particles. Budding then occurs, and release from the host cells is mediated by the neuraminidase activity of NA, which destroys the SA of the cellular and viral glycoproteins that would otherwise retain the new virions at the cell surface.
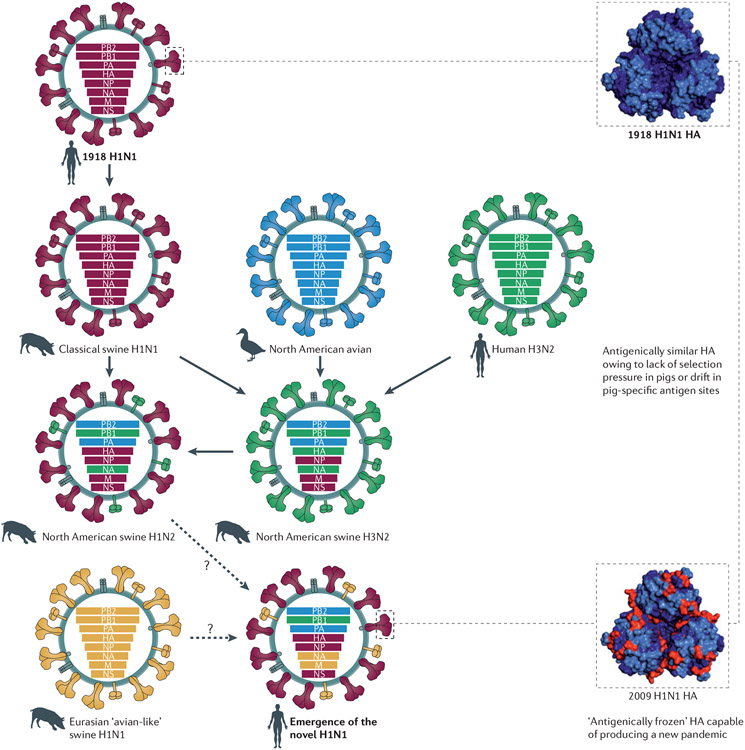
Influenza viruses similar to the 1918 pandemic H1N1 virus became established in domestic pigs between 1918 and 1920; this lineage is referred to as the classical swine lineage. In 1979, a distinct Eurasian ‘avian-like’ H1N1 virus emerged in European pigs and has since co-circulated with the classical swine H1N1 viruses. Triple-reassortant swine origin influenza virus (SOIV) H1 viruses of different strains and subtypes (for example, H3N2 and H1N2) emerged and became predominant among North American pig herds in the 1990s. All of these viruses provided the genetic pool for the genesis of the 2009 pandemic H1N1 SOIV, possibly owing to further reassortment in pigs. Thus, the 2009 pandemic H1N1 virus is composed of PB2 and PA segments from North American avian viruses, the PB1 segment of the human H3N2 viruses, haemagglutinin (HA; of the H1 subtype), nucleoprotein (NP) and NS segments derived from classical swine H1N1 viruses, and the neuraminidase (NA; of the N1 subtype) and M segments of Eurasian ‘avian-like’ swine viruses. Sequence and antigenic analyses of the 2009 pandemic H1N1 virus show that there are similarities between the HA of this virus and that of the 1918 and human H1N1 viruses that circulated sometime between 1918 and the 1950s. The antigenic similarities between the 1918 and 2009 pandemic H1N1 viruses are represented in the crystal structure models of the trimeric configuration of the HA protein globular head, as seen from a top view. The antigenic sites of the HA proteins are shown in light blue, non-antigenic sites are shown in dark blue. The sites that differ between the 1918 and 2009 HA proteins are depicted in red.
The anatomical expression patterns of the viral receptors in different hosts restricts infection with and replication of influenza A viruses. The swine trachea contains receptors with α-2,3-linked and α-2,6-linked sialic acid (α-2,3-SA and α-2,6-SA) moieties that allow for binding of both avian and human viruses, leading to the idea that pigs can serve as the ‘mixing vessel’ (REF 57) in which reassortment of human and avian viruses can occur. Avian viruses bind preferentially to α-2,3-SA, which is found on receptors in the gut and respiratory tract of birds. By contrast, human-adapted viruses (for example, seasonal H1N1, H3N2 and 2009 pandemic H1N1 viruses) have a higher affinity for α-2,6-SAs, which are expressed in the upper respiratory tract of humans. Human infection with a non-human-adapted virus is rare and is usually a result of a direct spillover transmission event. Viral proteins and their specific residues that affect receptor binding and have been established as adaptations to the human host are listed; H1, H3 and H5 are variations of the haemagglutinin (HA) protein, and PB2 is an RNA-dependent RNA polymerase component.

Virus–host interactions affect viral replication. Variation in many of the influenza A virus proteins contributes to pathogenesis, and host factors can also influence susceptibility to infection and disease progression. The risk factors obesity and diabetes were identified during the 2009 H1N1 pandemic. HA, haemagglutinin; NA, neuraminidase; NP, nucleoprotein; NS1, non-structural protein 1.
Similar articles
-
Virulence determinants of pandemic influenza viruses.J Clin Invest. 2011 Jan;121(1):6-13. doi: 10.1172/JCI44947. Epub 2011 Jan 4. J Clin Invest. 2011. PMID: 21206092 Free PMC article. Review.
-
Recent zoonoses caused by influenza A viruses.Rev Sci Tech. 2000 Apr;19(1):197-225. doi: 10.20506/rst.19.1.1220. Rev Sci Tech. 2000. PMID: 11189716 Review.
-
Influenza viruses: breaking all the rules.mBio. 2013 Jul 16;4(4):e00365-13. doi: 10.1128/mBio.00365-13. mBio. 2013. PMID: 23860766 Free PMC article.
-
[Interspecies transmission, adaptation to humans and pathogenicity of animal influenza viruses].Pathol Biol (Paris). 2010 Apr;58(2):e59-68. doi: 10.1016/j.patbio.2010.01.012. Epub 2010 Mar 19. Pathol Biol (Paris). 2010. PMID: 20303675 Review. French.
-
Predicting Zoonotic Risk of Influenza A Viruses from Host Tropism Protein Signature Using Random Forest.Int J Mol Sci. 2017 May 25;18(6):1135. doi: 10.3390/ijms18061135. Int J Mol Sci. 2017. PMID: 28587080 Free PMC article.
Cited by
-
CRISPR/Cas13a combined with hybridization chain reaction for visual detection of influenza A (H1N1) virus.Anal Bioanal Chem. 2022 Dec;414(29-30):8437-8445. doi: 10.1007/s00216-022-04380-1. Epub 2022 Oct 20. Anal Bioanal Chem. 2022. PMID: 36264297 Free PMC article.
-
An overview of the highly pathogenic H5N1 influenza virus.Virol Sin. 2013 Feb;28(1):3-15. doi: 10.1007/s12250-013-3294-9. Epub 2013 Jan 16. Virol Sin. 2013. PMID: 23325419 Free PMC article. Review.
-
Avian influenza A H5N1 virus: a continuous threat to humans.Emerg Microbes Infect. 2012 Sep;1(9):e25. doi: 10.1038/emi.2012.24. Epub 2012 Sep 19. Emerg Microbes Infect. 2012. PMID: 26038430 Free PMC article. Review.
-
Variable ligand- and receptor-binding hot spots in key strains of influenza neuraminidase.J Mol Genet Med. 2012;6:293-300. doi: 10.4172/1747-0862.1000052. Epub 2012 May 25. J Mol Genet Med. 2012. PMID: 22872804 Free PMC article.
-
Roles and functions of IAV proteins in host immune evasion.Front Immunol. 2023 Dec 13;14:1323560. doi: 10.3389/fimmu.2023.1323560. eCollection 2023. Front Immunol. 2023. PMID: 38152399 Free PMC article. Review.
References
-
- Molinari NA et al. The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine 25, 5086–5096 (2007). - PubMed
-
- Johnson NP & Mueller J Updating the accounts: global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull. Hist. Med 76, 105–115 (2002). - PubMed
-
- Palese P & Shaw ML in Fields Virology 5th edn (eds Knipe DM et al.) 1647–1689 (Lippincott Williams & Wilkins, Philadelphia, USA, 2007).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
