

Integrative regulatory mapping indicates that the RNA-binding protein HuR couples pre-mRNA processing and mRNA stability
- PMID: 21723170
- PMCID: PMC3220597
- DOI: 10.1016/j.molcel.2011.06.007
Integrative regulatory mapping indicates that the RNA-binding protein HuR couples pre-mRNA processing and mRNA stability
Abstract
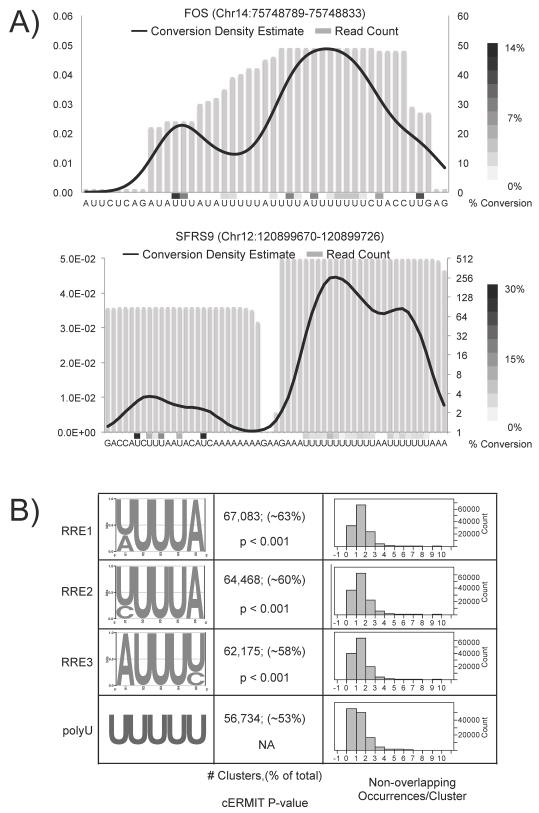
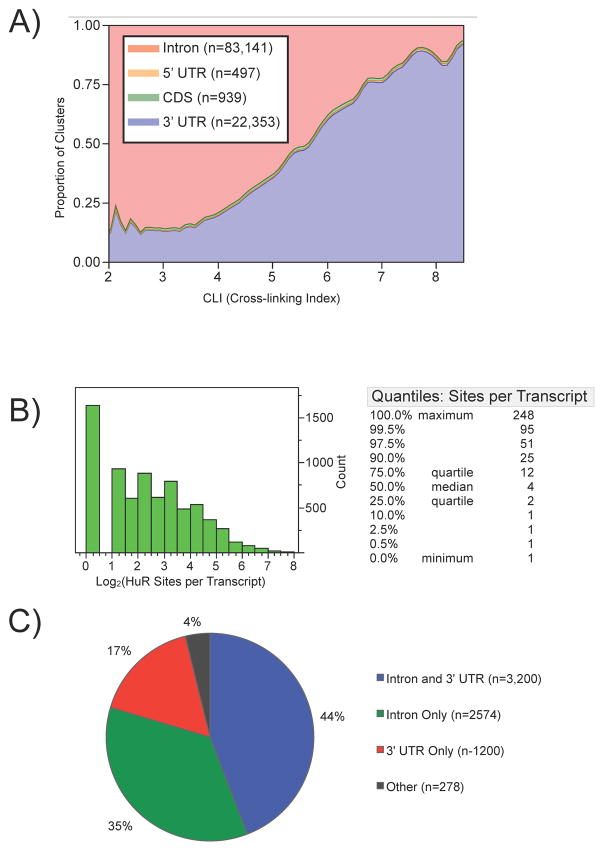
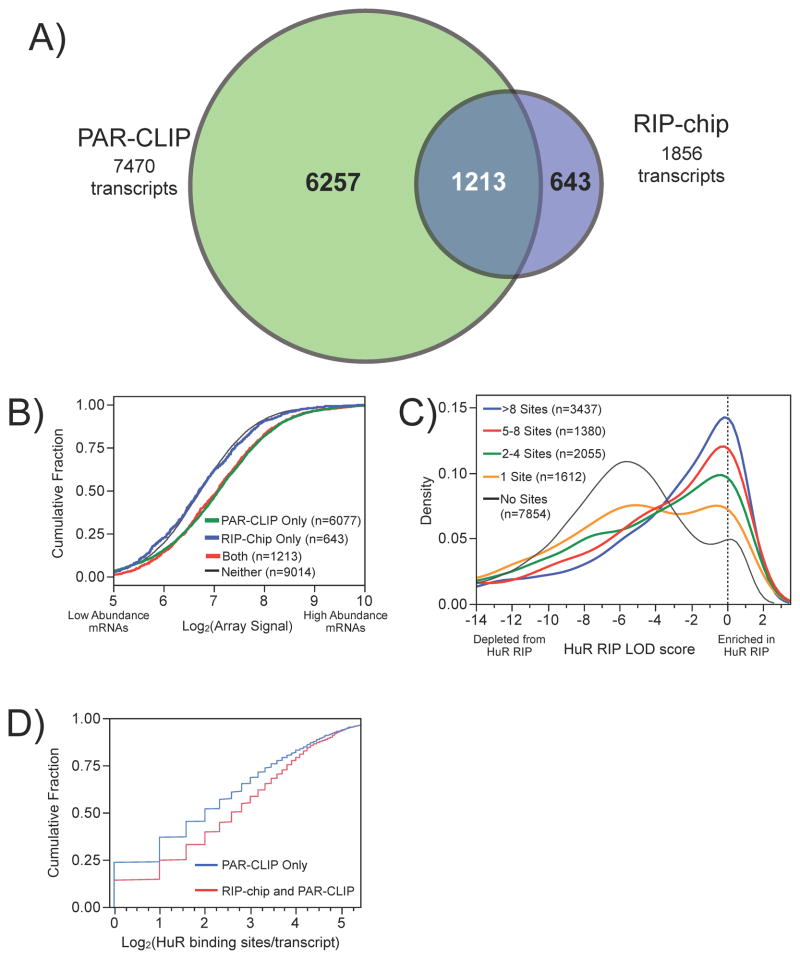
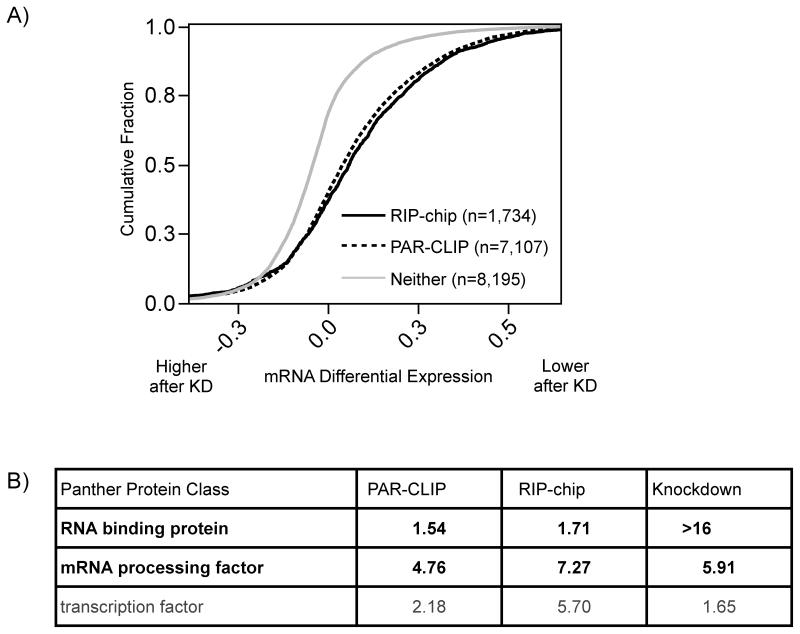
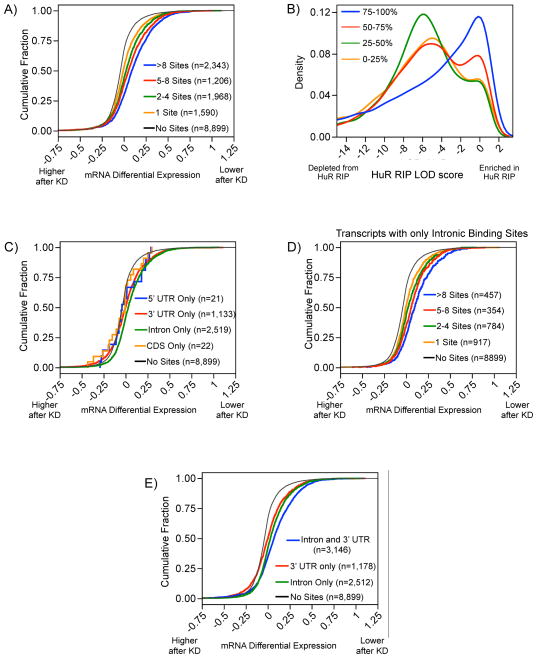
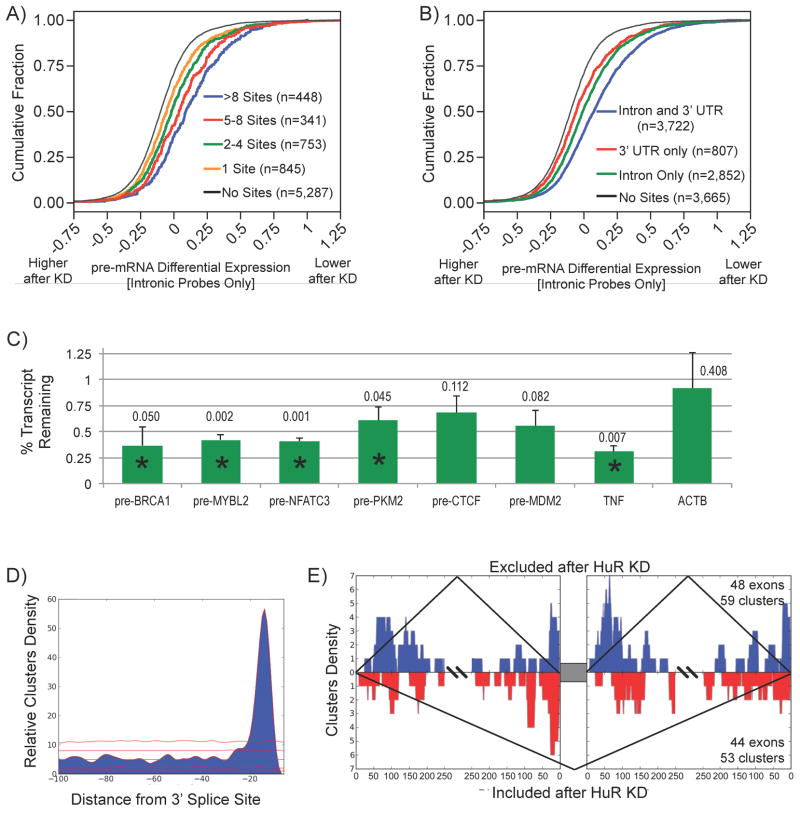
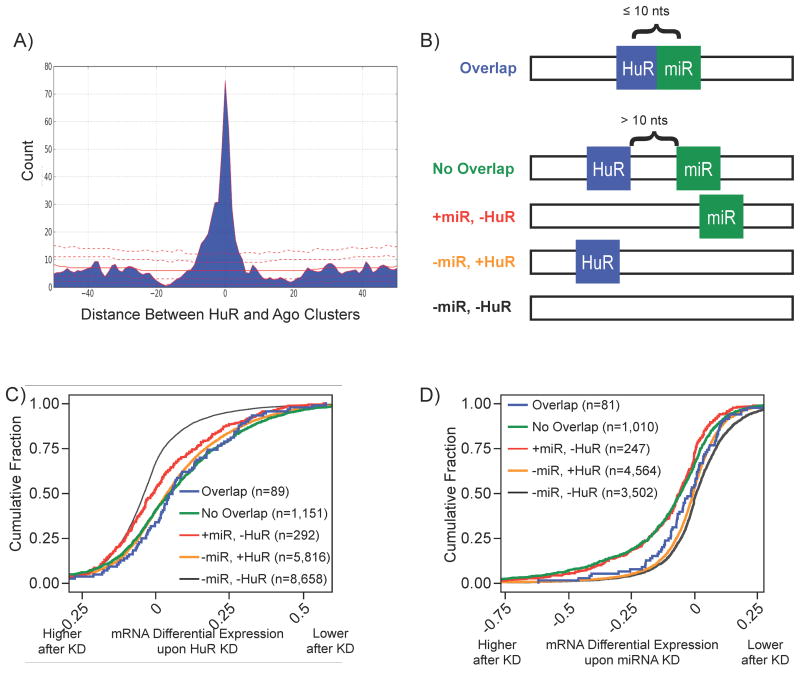
RNA-binding proteins coordinate the fates of multiple RNAs, but the principles underlying these global interactions remain poorly understood. We elucidated regulatory mechanisms of the RNA-binding protein HuR, by integrating data from diverse high-throughput targeting technologies, specifically PAR-CLIP, RIP-chip, and whole-transcript expression profiling. The number of binding sites per transcript, degree of HuR association, and degree of HuR-dependent RNA stabilization were positively correlated. Pre-mRNA and mature mRNA containing both intronic and 3' UTR binding sites were more highly stabilized than transcripts with only 3' UTR or only intronic binding sites, suggesting that HuR couples pre-mRNA processing with mature mRNA stability. We also observed HuR-dependent splicing changes and substantial binding of HuR in polypyrimidine tracts of pre-mRNAs. Comparison of the spatial patterns surrounding HuR and miRNA binding sites provided functional evidence for HuR-dependent antagonism of proximal miRNA-mediated repression. We conclude that HuR coordinates gene expression outcomes at multiple interconnected steps of RNA processing.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
UneCLIPsing HuR nuclear function.Mol Cell. 2011 Aug 5;43(3):319-21. doi: 10.1016/j.molcel.2011.07.016. Mol Cell. 2011. PMID: 21816340 Free PMC article.
Similar articles
-
Transcriptome-wide analysis of regulatory interactions of the RNA-binding protein HuR.Mol Cell. 2011 Aug 5;43(3):340-52. doi: 10.1016/j.molcel.2011.06.008. Epub 2011 Jun 30. Mol Cell. 2011. PMID: 21723171
-
Embryonic lethal abnormal vision-like HuR-dependent mRNA stability regulates post-transcriptional expression of cyclin-dependent kinase inhibitor p27Kip1.J Biol Chem. 2010 May 14;285(20):15408-15419. doi: 10.1074/jbc.M110.113365. Epub 2010 Mar 23. J Biol Chem. 2010. PMID: 20332085 Free PMC article.
-
RNPC1 modulates the RNA-binding activity of, and cooperates with, HuR to regulate p21 mRNA stability.Nucleic Acids Res. 2010 Apr;38(7):2256-67. doi: 10.1093/nar/gkp1229. Epub 2010 Jan 11. Nucleic Acids Res. 2010. PMID: 20064878 Free PMC article.
-
Properties of the regulatory RNA-binding protein HuR and its role in controlling miRNA repression.Adv Exp Med Biol. 2010;700:106-23. Adv Exp Med Biol. 2010. PMID: 21627034 Review.
-
HuR, a key post-transcriptional regulator, and its implication in progression of breast cancer.Histol Histopathol. 2010 Oct;25(10):1331-40. doi: 10.14670/HH-25.1331. Histol Histopathol. 2010. PMID: 20712017 Review.
Cited by
-
Intronic RNA secondary structural information captured for the human MYC pre-mRNA.NAR Genom Bioinform. 2024 Oct 24;6(4):lqae143. doi: 10.1093/nargab/lqae143. eCollection 2024 Sep. NAR Genom Bioinform. 2024. PMID: 39450312 Free PMC article.
-
LincRNA-p21 suppresses target mRNA translation.Mol Cell. 2012 Aug 24;47(4):648-55. doi: 10.1016/j.molcel.2012.06.027. Epub 2012 Jul 26. Mol Cell. 2012. PMID: 22841487 Free PMC article.
-
Mechanisms coordinating ELAV/Hu mRNA regulons.Curr Opin Genet Dev. 2013 Feb;23(1):35-43. doi: 10.1016/j.gde.2012.12.006. Epub 2013 Jan 9. Curr Opin Genet Dev. 2013. PMID: 23312841 Free PMC article. Review.
-
Hsp70 is a novel posttranscriptional regulator of gene expression that binds and stabilizes selected mRNAs containing AU-rich elements.Mol Cell Biol. 2013 Jan;33(1):71-84. doi: 10.1128/MCB.01275-12. Epub 2012 Oct 29. Mol Cell Biol. 2013. PMID: 23109422 Free PMC article.
-
mRNA fate: Life and death of the mRNA in the cytoplasm.RNA Biol. 2013 Mar;10(3):360-6. doi: 10.4161/rna.23770. Epub 2013 Mar 1. RNA Biol. 2013. PMID: 23466755 Free PMC article.
References
-
- Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W. Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell. 2006;125:1111–1124. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
