

Apical deficiency triggers JNK-dependent apoptosis in the embryonic epidermis of Drosophila
- PMID: 21693518
- PMCID: PMC3119309
- DOI: 10.1242/dev.059980
Apical deficiency triggers JNK-dependent apoptosis in the embryonic epidermis of Drosophila
Abstract
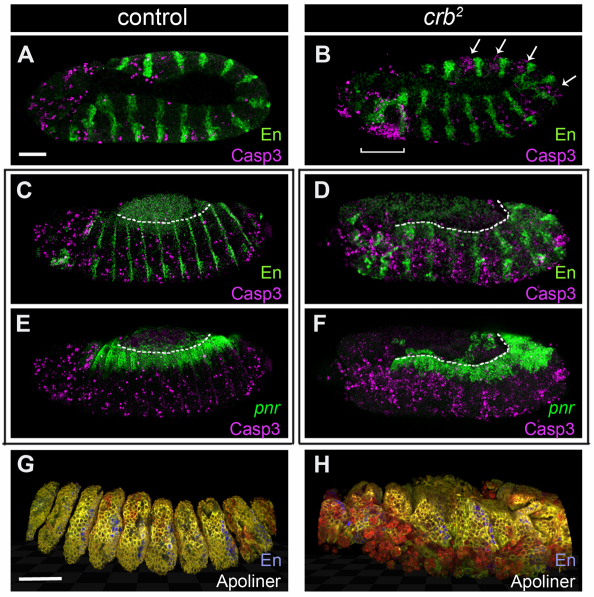
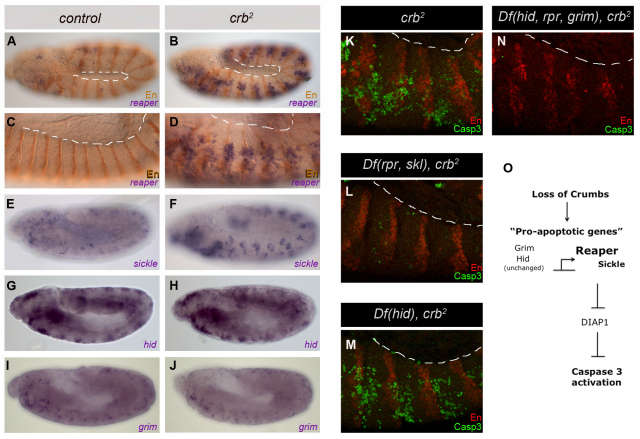
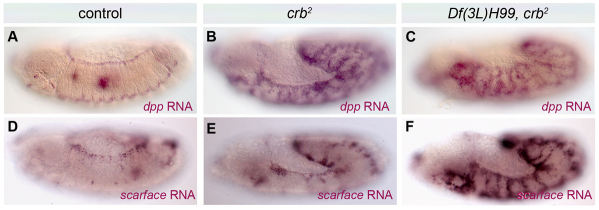
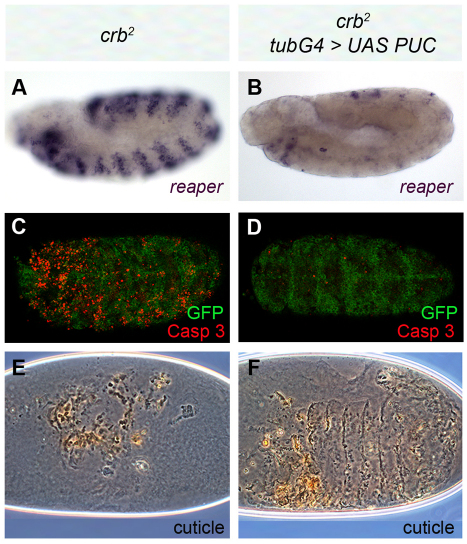
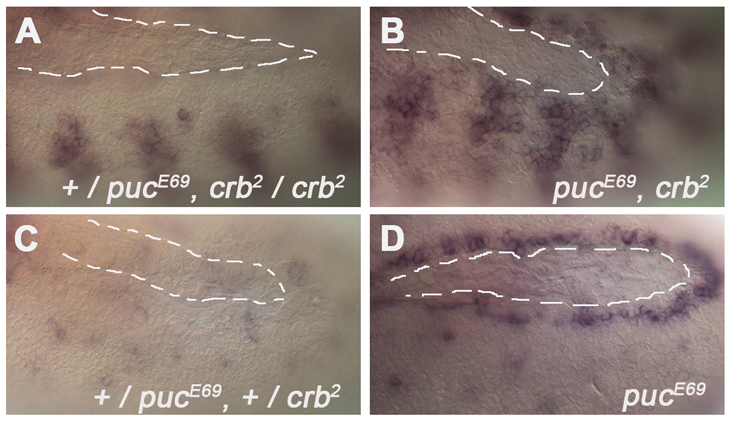
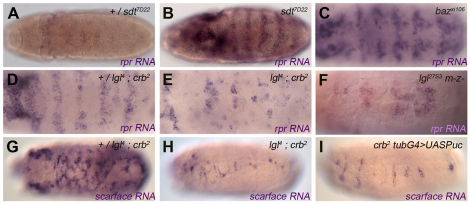
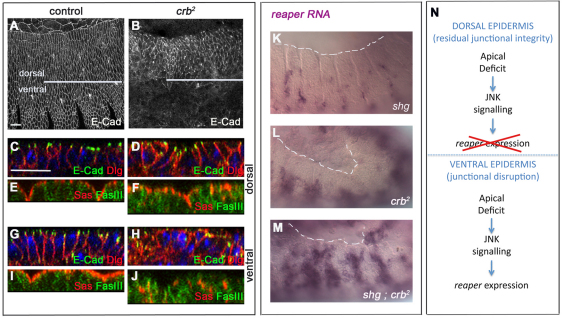
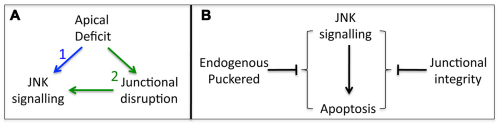
Epithelial homeostasis and the avoidance of diseases such as cancer require the elimination of defective cells by apoptosis. Here, we investigate how loss of apical determinants triggers apoptosis in the embryonic epidermis of Drosophila. Transcriptional profiling and in situ hybridisation show that JNK signalling is upregulated in mutants lacking Crumbs or other apical determinants. This leads to transcriptional activation of the pro-apoptotic gene reaper and to apoptosis. Suppression of JNK signalling by overexpression of Puckered, a feedback inhibitor of the pathway, prevents reaper upregulation and apoptosis. Moreover, removal of endogenous Puckered leads to ectopic reaper expression. Importantly, disruption of the basolateral domain in the embryonic epidermis does not trigger JNK signalling or apoptosis. We suggest that apical, not basolateral, integrity could be intrinsically required for the survival of epithelial cells. In apically deficient embryos, JNK signalling is activated throughout the epidermis. Yet, in the dorsal region, reaper expression is not activated and cells survive. One characteristic of these surviving cells is that they retain discernible adherens junctions despite the apical deficit. We suggest that junctional integrity could restrain the pro-apoptotic influence of JNK signalling.
Figures
Similar articles
-
Puckered, a Drosophila MAPK phosphatase, ensures cell viability by antagonizing JNK-induced apoptosis.Development. 2005 Sep;132(17):3935-46. doi: 10.1242/dev.01949. Epub 2005 Aug 3. Development. 2005. PMID: 16079158
-
Drosophila jumu modulates apoptosis via a JNK-dependent pathway and is required for other processes in wing development.Apoptosis. 2019 Jun;24(5-6):465-477. doi: 10.1007/s10495-019-01527-x. Apoptosis. 2019. PMID: 30796611
-
Spoonbill positively regulates JNK signalling mediated apoptosis in Drosophila melanogaster.Eur J Cell Biol. 2023 Jun;102(2):151300. doi: 10.1016/j.ejcb.2023.151300. Epub 2023 Feb 24. Eur J Cell Biol. 2023. PMID: 36858008
-
The Dpp/TGFβ-dependent corepressor Schnurri protects epithelial cells from JNK-induced apoptosis in drosophila embryos.Dev Cell. 2014 Oct 27;31(2):240-7. doi: 10.1016/j.devcel.2014.08.015. Epub 2014 Oct 9. Dev Cell. 2014. PMID: 25307481 Free PMC article.
-
Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila.Differentiation. 2002 Jun;70(4-5):181-203. doi: 10.1046/j.1432-0436.2002.700408.x. Differentiation. 2002. PMID: 12147138 Review.
Cited by
-
Apical polarity proteins recruit the RhoGEF Cysts to promote junctional myosin assembly.J Cell Biol. 2019 Oct 7;218(10):3397-3414. doi: 10.1083/jcb.201807106. Epub 2019 Aug 13. J Cell Biol. 2019. PMID: 31409654 Free PMC article.
-
Rab11 negatively regulates wingless preventing JNK-mediated apoptosis in Drosophila epithelium during embryonic dorsal closure.Cell Tissue Res. 2023 Mar;391(3):485-504. doi: 10.1007/s00441-023-03740-2. Epub 2023 Jan 27. Cell Tissue Res. 2023. PMID: 36705747
-
Epithelial cells release adenosine to promote local TNF production in response to polarity disruption.Nat Commun. 2018 Nov 7;9(1):4675. doi: 10.1038/s41467-018-07114-z. Nat Commun. 2018. PMID: 30405122 Free PMC article.
-
A dual role of the extracellular domain of Drosophila Crumbs for morphogenesis of the embryonic neuroectoderm.Biol Open. 2018 Jan 26;7(1):bio031435. doi: 10.1242/bio.031435. Biol Open. 2018. PMID: 29374056 Free PMC article.
-
Vasopressin decreases neuronal apoptosis during cardiopulmonary resuscitation.Neural Regen Res. 2014 Mar 15;9(6):622-9. doi: 10.4103/1673-5374.130107. Neural Regen Res. 2014. PMID: 25206865 Free PMC article.
References
-
- Abrams J. M., White K., Fessler L. I., Steller H. (1993). Programmed cell death during Drosophila embryogenesis. Development 117, 29-43 - PubMed
-
- Adachi-Yamada T., O'Connor M. B. (2002). Morphogenetic apoptosis: a mechanism for correcting discontinuities in morphogen gradients. Dev. Biol. 251, 74-90 - PubMed
-
- Affolter M., Nellen D., Nussbaumer U., Basler K. (1994). Multiple requirements for the receptor serine/threonine kinase thick veins reveal novel functions of TGF beta homologs during Drosophila embryogenesis. Development 120, 3105-3117 - PubMed
-
- Agrawal N., Kango M., Mishra A., Sinha P. (1995). Neoplastic transformation and aberrant cell-cell interactions in genetic mosaics of lethal(2)giant larvae (lgl), a tumor suppressor gene of Drosophila. Dev. Biol. 172, 218-229 - PubMed
-
- Baehrecke E. H. (2002). How death shapes life during development. Nat. Rev. Mol. Cell Biol. 3, 779-787 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
