

Roles of the fusion and hemagglutinin-neuraminidase proteins in replication, tropism, and pathogenicity of avian paramyxoviruses
- PMID: 21680512
- PMCID: PMC3165810
- DOI: 10.1128/JVI.00652-11
Roles of the fusion and hemagglutinin-neuraminidase proteins in replication, tropism, and pathogenicity of avian paramyxoviruses
Abstract
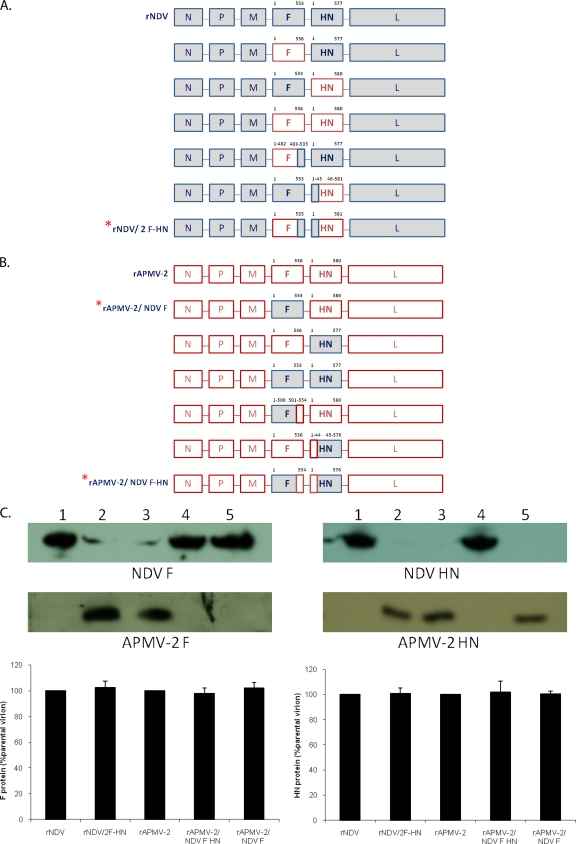
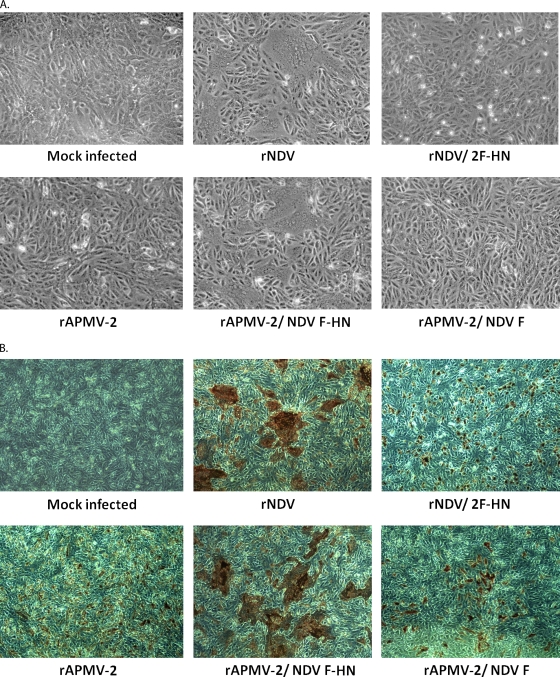
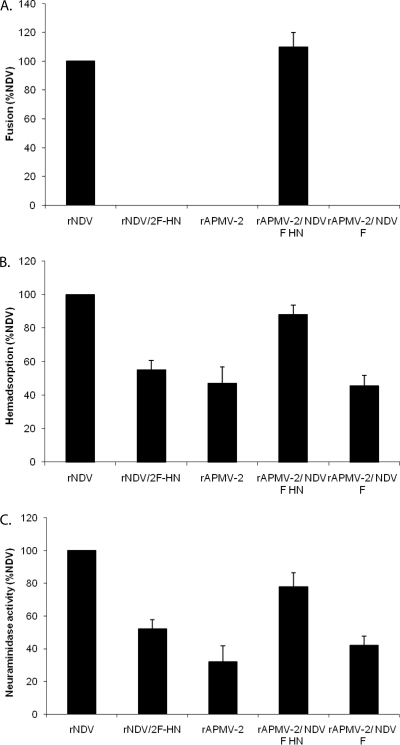
Virulent and moderately virulent strains of Newcastle disease virus (NDV), representing avian paramyxovirus serotype 1 (APMV-1), cause respiratory and neurological disease in chickens and other species of birds. In contrast, APMV-2 is avirulent in chickens. We investigated the role of the fusion (F) and hemagglutinin-neuraminidase (HN) envelope glycoproteins in these contrasting phenotypes by designing chimeric viruses in which the F and HN glycoproteins or their ectodomains were exchanged individually or together between the moderately virulent, neurotropic NDV strain Beaudette C (BC) and the avirulent APMV-2 strain Yucaipa. When we attempted to exchange the complete F and HN glycoproteins individually and together between the two viruses, the only construct that could be recovered was recombinant APMV-2 strain Yucaipa (rAPMV-2), containing the NDV F glycoprotein in place of its own. This substitution of NDV F into APMV-2 was sufficient to confer the neurotropic, neuroinvasive, and neurovirulent phenotypes, in spite of all being at reduced levels compared to what was seen for NDV-BC. When the ectodomains of F and HN were exchanged individually and together, two constructs could be recovered: NDV, containing both the F and HN ectodomains of APMV-2; and APMV-2, containing both ectodomains of NDV. This supported the idea that homologous cytoplasmic tails and matched F and HN ectodomains are important for virus replication. Analysis of these viruses for replication in vitro, syncytium formation, mean embryo death time, intracerebral pathogenicity index, and replication and tropism in 1-day-old chicks and 2-week-old chickens showed that the two contrasting phenotypes of NDV and APMV-2 could largely be transferred between the two backbones by transfer of homotypic F and HN ectodomains. Further analysis provided evidence that the homologous stalk domain of NDV HN is essential for virus replication, while the globular head domain of NDV HN could be replaced with that of APMV-2 with only a minimal attenuating effect. These results demonstrate that the F and HN ectodomains together determine the cell fusion, tropism, and virulence phenotypes of NDV and APMV-2 and that the regions of HN that are critical to replication and the species-specific phenotypes include the cytoplasmic tail and stalk domain but not the globular head domain.
Figures

Similar articles
-
Evaluation of the Newcastle disease virus F and HN proteins in protective immunity by using a recombinant avian paramyxovirus type 3 vector in chickens.J Virol. 2011 Jul;85(13):6521-34. doi: 10.1128/JVI.00367-11. Epub 2011 Apr 27. J Virol. 2011. Retraction in: J Virol. 2020 Feb 28;94(6):e01867-19. doi: 10.1128/JVI.01867-19 PMID: 21525340 Free PMC article. Retracted.
-
Generation of a recombinant chimeric Newcastle disease virus vaccine that allows serological differentiation between vaccinated and infected animals.Vaccine. 2001 Feb 8;19(13-14):1616-27. doi: 10.1016/s0264-410x(00)00419-9. Vaccine. 2001. PMID: 11166884
-
Newcastle Disease Virus Establishes Persistent Infection in Tumor Cells In Vitro: Contribution of the Cleavage Site of Fusion Protein and Second Sialic Acid Binding Site of Hemagglutinin-Neuraminidase.J Virol. 2017 Jul 27;91(16):e00770-17. doi: 10.1128/JVI.00770-17. Print 2017 Aug 15. J Virol. 2017. PMID: 28592535 Free PMC article.
-
Molecular characterization of hemagglutinin-neuraminidase fragment gene of Newcastle disease virus isolated from periodically-vaccinated farms.Vet World. 2018 May;11(5):657-666. doi: 10.14202/vetworld.2018.657-666. Epub 2018 May 20. Vet World. 2018. PMID: 29915505 Free PMC article. Review.
-
Multifunctionality of matrix protein in the replication and pathogenesis of Newcastle disease virus: A review.Int J Biol Macromol. 2023 Sep 30;249:126089. doi: 10.1016/j.ijbiomac.2023.126089. Epub 2023 Jul 31. Int J Biol Macromol. 2023. PMID: 37532184 Review.
Cited by
-
Comparative Protective Efficacies of Novel Avian Paramyxovirus-Vectored Vaccines against Virulent Infectious Bronchitis Virus in Chickens.Viruses. 2020 Jun 28;12(7):697. doi: 10.3390/v12070697. Viruses. 2020. PMID: 32605292 Free PMC article.
-
Identification of a new amino acid mutation in the HN protein of NDV involved in pathogenicity.Vet Res. 2021 Dec 20;52(1):147. doi: 10.1186/s13567-021-01019-4. Vet Res. 2021. PMID: 34930432 Free PMC article.
-
Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya.Viruses. 2021 Jan 13;13(1):103. doi: 10.3390/v13010103. Viruses. 2021. PMID: 33451125 Free PMC article.
-
Complete genome sequence of a novel avian paramyxovirus.J Virol. 2012 Jul;86(14):7710. doi: 10.1128/JVI.00946-12. J Virol. 2012. PMID: 22733876 Free PMC article.
-
Functional and structural analyses reveal that a dual domain sialidase protects bacteria from complement killing through desialylation of complement factors.PLoS Pathog. 2023 Sep 25;19(9):e1011674. doi: 10.1371/journal.ppat.1011674. eCollection 2023 Sep. PLoS Pathog. 2023. PMID: 37747935 Free PMC article.
References
-
- Alexander D. J. 1982. Avian paramyxoviruses-other than Newcastle disease virus. Worlds Poult. Sci. J. 38:97–104
-
- Alexander D. J. 1989. Newcastle disease, p. 114–120 In Purchase H. G., Arp L. H., Domermuth C. H., Pearson J. E. (ed.), A laboratory manual for the isolation and identification of avian pathogens, 3rd ed The American Association of Avian Pathologists, Kendall/Hunt Publishing Company, Dubuque, IA
-
- Alexander D. J. 2003. Avian paramyxoviruses 2–9, p. 88–92 In Saif Y. M. (ed.), Diseases of poultry, 11th ed Iowa State University Press, Ames, IA
-
- de Leeuw O. S., Koch G., Hartog L., Ravenshorst N., Peeters B. P. 2005. Virulence of Newcastle disease virus is determined by the cleavage site of the fusion protein and by both the stem region and globular head of the haemagglutinin-neuraminidase protein. J. Gen. Virol. 86:1759–1769 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
