

Review
doi: 10.1101/cshperspect.a007559.
Quality control of mitochondrial proteostasis
Affiliations
- PMID: 21628427
- PMCID: PMC3119916
- DOI: 10.1101/cshperspect.a007559
Item in Clipboard
Review
Quality control of mitochondrial proteostasis
Cold Spring Harb Perspect Biol.
.
Abstract
A decline in mitochondrial activity has been associated with aging and is a hallmark of many neurological diseases. Surveillance mechanisms acting at the molecular, organellar, and cellular level monitor mitochondrial integrity and ensure the maintenance of mitochondrial proteostasis. Here we will review the central role of mitochondrial chaperones and proteases, the cytosolic ubiquitin-proteasome system, and the mitochondrial unfolded response in this interconnected quality control network, highlighting the dual function of some proteases in protein quality control within the organelle and for the regulation of mitochondrial fusion and mitophagy.
Figures

Mitochondrial surveillance mechanisms. Low levels of mitochondrial damage are dealt with on the molecular level by conserved proteolytic machineries. Proteins that are unfolded may be refolded by chaperones or degraded by proteases. Mitochondrial dynamics contributes to the functional integrity of mitochondria. Mitochondrial fusion permits content mixing, and stress-induced mitochondrial hyperfusion (SIMH) may alleviate mild mitochondrial stress. Severe mitochondrial damage or depolarization of the mitochondrial membrane potential inhibits fusion, and ongoing fission will segregate the damaged mitochondria from the healthy network, allowing its turnover through mitophagy. Finally, if the level of damaged mitochondria becomes too high and mitochondrial outer membrane permeablization (MOMP) takes place, cytochrome c is released initiating cell death and thus turnover of the entire cell.
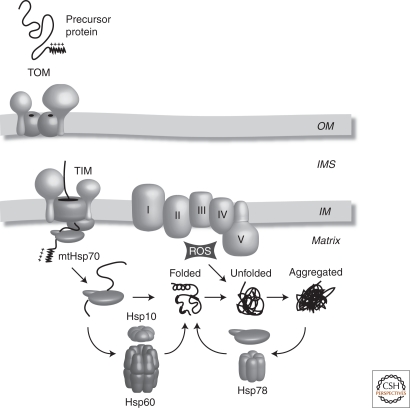
Chaperone systems in the mitochondrial matrix. mtHsp70 drives the import of nuclear encoded preproteins through the TOM and TIM (translocases in the OM and IM, respectively). In addition, mtHsp70 facilitates protein folding, although some proteins require an additional chaperonin complex, Hsp60/10, to fold. Stress-induced unfolding renders proteins prone to aggregation. The Hsp78 chaperone acts as a disaggregase and in conjunction with mtHsp70 can restore a proteins tertiary structure. OM, mitochondrial outer membrane; IMS, mitochondrial intermembrane space; IM, mitochondrial inner membrane; I–V, respiratory chain complexes, ROS, reactive oxygen species.
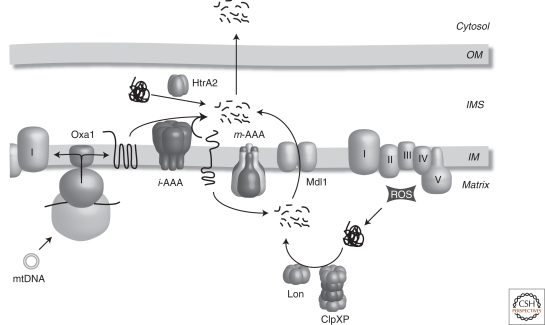
Proteolytic systems in mitochondria. Newly synthesized subunits of the mitochondrial respiratory chain are inserted into the IM by Oxa1 and assemble with newly imported nuclear encoded subunits. Unassembled subunits as well as unfolded or damaged IM proteins are turned over by membrane embedded AAA protease complexes. Proteins that become damaged or unfolded in the mitochondrial matrix are degraded by Lon or ClpXP (in higher eukaryotes). Peptides that are generated in the mitochondrial matrix are exported from matrix through the peptide transporter Mdl1 and eventually out of the organelle. HtrA2 may play a role in turnover of misfolded or unassembled protein in the IMS. OM, mitochondrial outer membrane; IMS, mitochondrial intermembrane space; IM, mitochondrial inner membrane; I–V, respiratory chain complexes; ROS, reactive oxygen species.
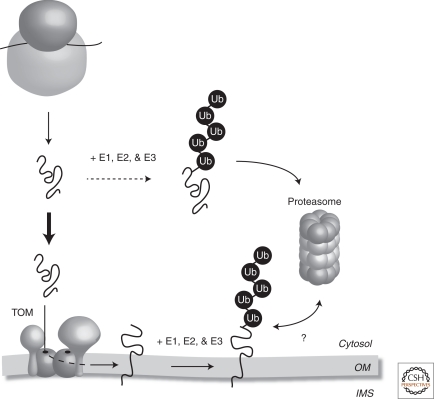
Mitochondrial proteins and the ubiquitin-proteasome system. Following their synthesis in the cytosol, mitochondrial precursor proteins follow two pathways. The first (bold line) ensures that the precursor protein is efficiently targeted to the translocase of the outer membrane (TOM). Some proteins that reside in the mitochondrial OM can be ubiquitinated (Ub) by a cascade of E1, E2, and E3 enzymes and subsequently turned over in an ubiquitin-proteasome-dependent pathway. It remains unclear if mitochondrial OM proteins are first extracted from the OM and delivered to the proteasome (indicated by the ?). It is possible that components are recruited to the mitochondrial surface to extract the protein or that the proteasome itself is recruited to the mitochondrial surface to facilitate protein turnover. The second pathway (dashed line) dictates that a fraction of newly synthesized mitochondrial precursor proteins will be ubiquitinated and degraded by the 26S proteasome in the cytosol before reaching the mitochondria. Precursor proteins that contain mutations or have been mistargeted may also be degraded in the cytosol. OM, mitochondrial outer membrane; IMS, mitochondrial intermembrane space.
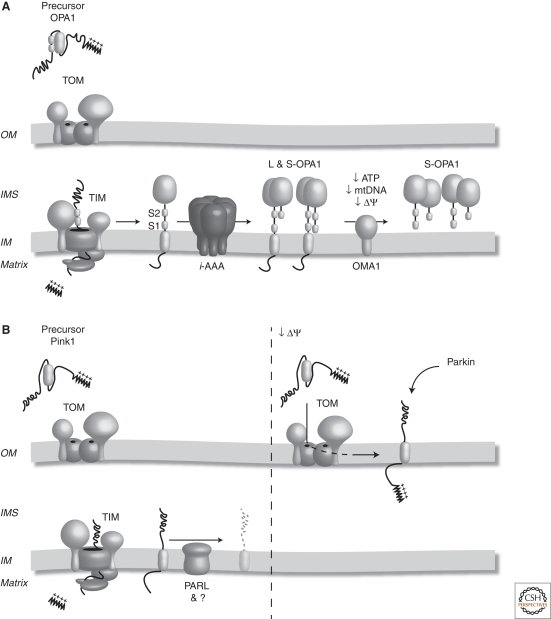
Mitochondrial proteases influence mitochondrial morphology and turnover. (A) OPA1 is synthesized in the cytosol and targeted to mitochondria via its amino-terminal mitochondrial targeting sequence. Following import into the organelle, the targeting signal is cleaved and the hydrophobic transmembrane domain anchors OPA1 in the IM. Following lateral release into the lipid bilayer, OPA1 is subjected to constitutive processing at site 1 (S1) and by Yme1 at site 2 (S2), generating long and short forms of the protein (L and S OPA1) that ensure mitochondrial fusion. Under conditions of mitochondrial dysfunction, including decreased ATP, mtDNA, or membrane potential (ΔΨ), the long OPA1 isoforms are degraded by OMA1, resulting in mitochondrial fragmentation. (B) Pink1 is synthesized in the cytosol with a cleavable mitochondrial targeting sequence. Following its import into the IM, the mitochondrial targeting sequence is cleaved. Pink1 is then processed by PARL and turned over. Under conditions of mitochondrial dysfunction, such as a decrease in membrane potential (ΔΨ), Pink1 is alternately sorted to the mitochondrial OM where it can recruit Parkin to the mitochondrial surface and trigger mitophagy.
Similar articles
-
Quality control of the mitochondrion.Dev Cell. 2021 Apr 5;56(7):881-905. doi: 10.1016/j.devcel.2021.02.009. Epub 2021 Mar 3. Dev Cell. 2021. PMID: 33662258 Review.
-
Mitochondria-Associated Degradation Pathway (MAD) Function beyond the Outer Membrane.Cell Rep. 2020 Jul 14;32(2):107902. doi: 10.1016/j.celrep.2020.107902. Cell Rep. 2020. PMID: 32668258 Free PMC article.
-
Control of mitochondrial biogenesis and function by the ubiquitin-proteasome system.Open Biol. 2017 Apr;7(4):170007. doi: 10.1098/rsob.170007. Open Biol. 2017. PMID: 28446709 Free PMC article. Review.
-
Inhibition of proteasome reveals basal mitochondrial ubiquitination.J Proteomics. 2020 Oct 30;229:103949. doi: 10.1016/j.jprot.2020.103949. Epub 2020 Aug 31. J Proteomics. 2020. PMID: 32882436
-
Mitochondrial protein homeostasis: the cooperative roles of chaperones and proteases.Res Microbiol. 2009 Nov;160(9):718-25. doi: 10.1016/j.resmic.2009.08.003. Epub 2009 Aug 31. Res Microbiol. 2009. PMID: 19723579 Review.
Cited by
-
Mitochondrial fission, fusion, and stress.Science. 2012 Aug 31;337(6098):1062-5. doi: 10.1126/science.1219855. Science. 2012. PMID: 22936770 Free PMC article. Review.
-
Small heat shock proteins operate as molecular chaperones in the mitochondrial intermembrane space.Nat Cell Biol. 2023 Mar;25(3):467-480. doi: 10.1038/s41556-022-01074-9. Epub 2023 Jan 23. Nat Cell Biol. 2023. PMID: 36690850 Free PMC article.
-
Bioabsorbable metal zinc differentially affects mitochondria in vascular endothelial and smooth muscle cells.Biomater Biosyst. 2021 Aug 26;4:100027. doi: 10.1016/j.bbiosy.2021.100027. eCollection 2021 Dec. Biomater Biosyst. 2021. PMID: 36824572 Free PMC article.
-
The different axes of the mammalian mitochondrial unfolded protein response.BMC Biol. 2018 Jul 26;16(1):81. doi: 10.1186/s12915-018-0548-x. BMC Biol. 2018. PMID: 30049264 Free PMC article. Review.
-
Peroxisome Proliferator-Activated Receptor α Attenuates Hypertensive Vascular Remodeling by Protecting Vascular Smooth Muscle Cells from Angiotensin II-Induced ROS Production.Antioxidants (Basel). 2022 Nov 30;11(12):2378. doi: 10.3390/antiox11122378. Antioxidants (Basel). 2022. PMID: 36552585 Free PMC article.
References
-
- Anesti V, Scorrano L 2006. The relationship between mitochondrial shape and function and the cytoskeleton. Biochim Biophys Acta 1757: 692–699 - PubMed
-
- Arlt H, Tauer R, Feldmann H, Neupert W, Langer T 1996. The YTA10-12 complex, an AAA protease with chaperone-like activity in the inner membrane of mitochondria. Cell 85: 875–885 - PubMed
-
- Arnold I, Wagner-Ecker M, Ansorge W, Langer T 2006. Evidence for a novel mitochondria-to-nucleus signalling pathway in respiring cells lacking i-AAA protease and the ABC-transporter Mdl1. Gene 367: 74–88 - PubMed
-
- Augustin S, Nolden M, Muller S, Hardt O, Arnold I, Langer T 2005. Characterization of peptides released from mitochondria: Evidence for constant proteolysis and peptide efflux. J Biol Chem 280: 2691–2699 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources