

Escherichia coli Fpg glycosylase is nonrendundant and required for the rapid global repair of oxidized purine and pyrimidine damage in vivo
- PMID: 21601577
- PMCID: PMC3156590
- DOI: 10.1016/j.jmb.2011.05.004
Escherichia coli Fpg glycosylase is nonrendundant and required for the rapid global repair of oxidized purine and pyrimidine damage in vivo
Abstract
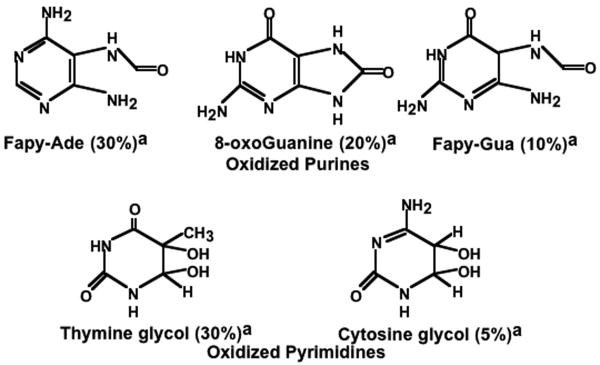
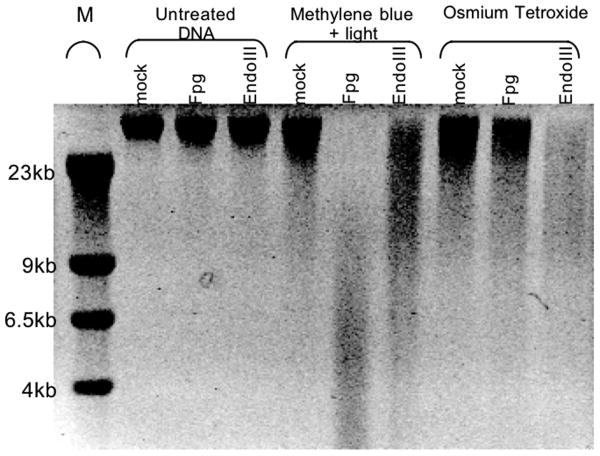
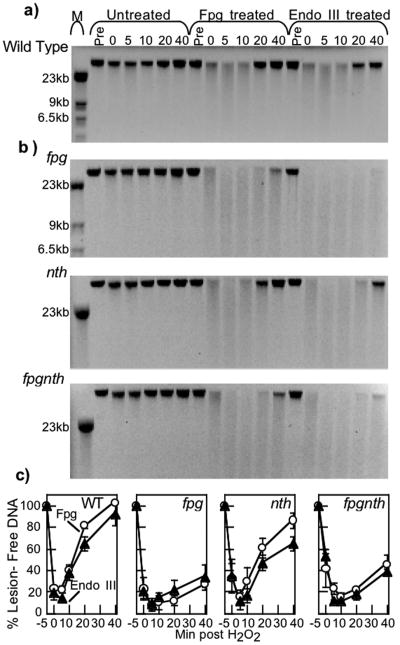
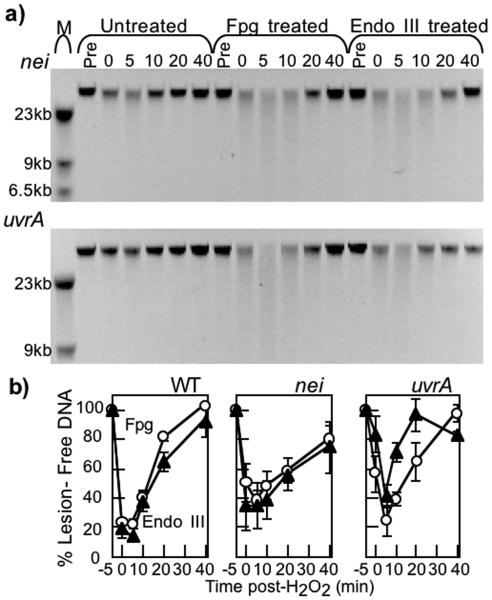
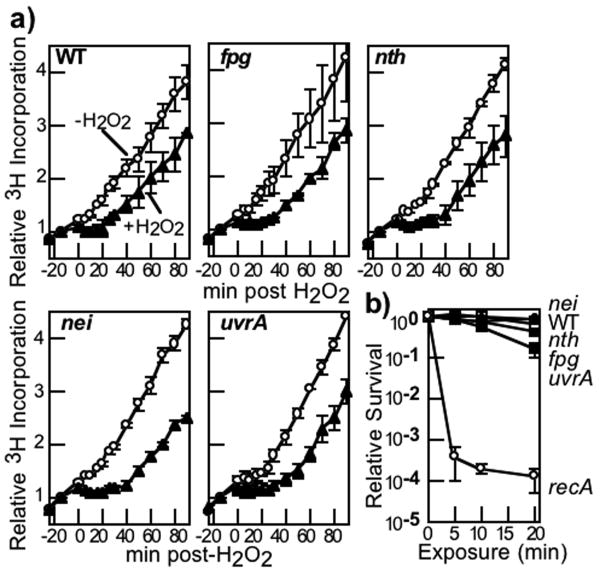
Endonuclease (Endo) III and formamidopyrimidine-N-glycosylase (Fpg) are two of the predominant DNA glycosylases in Escherichia coli that remove oxidative base damage. In cell extracts and purified form, Endo III is generally more active toward oxidized pyrimidines, while Fpg is more active towards oxidized purines. However, the substrate specificities of these enzymes partially overlap in vitro. Less is known about the relative contribution of these enzymes in restoring the genomic template following oxidative damage. In this study, we examined how efficiently Endo III and Fpg repair their oxidative substrates in vivo following treatment with hydrogen peroxide. We found that Fpg was nonredundant and required to rapidly remove its substrate lesions on the chromosome. In addition, Fpg also repaired a significant portion of the lesions recognized by Endo III, suggesting that it plays a prominent role in the global repair of both purine damage and pyrimidine damage in vivo. By comparison, Endo III did not affect the repair rate of Fpg substrates and was only responsible for repairing a subset of its own substrate lesions in vivo. The absence of Endo VIII or nucleotide excision repair did not significantly affect the global repair of either Fpg or Endo III substrates in vivo. Surprisingly, replication recovered after oxidative DNA damage in all mutants examined, even when lesions persisted in the DNA, suggesting the presence of an efficient mechanism to process or overcome oxidative damage encountered during replication.
Published by Elsevier Ltd.
Figures
Similar articles
-
DNA substrates containing defined oxidative base lesions and their application to study substrate specificities of base excision repair enzymes.Prog Nucleic Acid Res Mol Biol. 2001;68:207-21. doi: 10.1016/s0079-6603(01)68101-7. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11554298 Review.
-
The Fpg/Nei family of DNA glycosylases: substrates, structures, and search for damage.Prog Mol Biol Transl Sci. 2012;110:71-91. doi: 10.1016/B978-0-12-387665-2.00004-3. Prog Mol Biol Transl Sci. 2012. PMID: 22749143 Free PMC article. Review.
-
Excision of formamidopyrimidine lesions by endonucleases III and VIII is not a major DNA repair pathway in Escherichia coli.Nucleic Acids Res. 2005 Jun 8;33(10):3331-8. doi: 10.1093/nar/gki655. Print 2005. Nucleic Acids Res. 2005. PMID: 15944451 Free PMC article.
-
A novel role for Escherichia coli endonuclease VIII in prevention of spontaneous G-->T transversions.J Bacteriol. 1999 Oct;181(20):6396-402. doi: 10.1128/JB.181.20.6396-6402.1999. J Bacteriol. 1999. PMID: 10515930 Free PMC article.
-
Excision of the oxidatively formed 5-hydroxyhydantoin and 5-hydroxy-5-methylhydantoin pyrimidine lesions by Escherichia coli and Saccharomyces cerevisiae DNA N-glycosylases.Biochim Biophys Acta. 2009 Jan 1;1790(1):16-24. doi: 10.1016/j.bbagen.2008.10.001. Epub 2008 Oct 17. Biochim Biophys Acta. 2009. PMID: 18983898
Cited by
-
Evaluation of oxidative stress responses in human circulating blood cells after imatinib mesylate treatment - Implications to its mechanism of action.Saudi Pharm J. 2019 Dec;27(8):1216-1221. doi: 10.1016/j.jsps.2019.10.005. Epub 2019 Oct 25. Saudi Pharm J. 2019. PMID: 31885482 Free PMC article.
-
Identification of genes involved in low aminoglycoside-induced SOS response in Vibrio cholerae: a role for transcription stalling and Mfd helicase.Nucleic Acids Res. 2014 Feb;42(4):2366-79. doi: 10.1093/nar/gkt1259. Epub 2013 Dec 6. Nucleic Acids Res. 2014. PMID: 24319148 Free PMC article.
-
Oxidative DNA damage is instrumental in hyperreplication stress-induced inviability of Escherichia coli.Nucleic Acids Res. 2014 Dec 1;42(21):13228-41. doi: 10.1093/nar/gku1149. Epub 2014 Nov 11. Nucleic Acids Res. 2014. PMID: 25389264 Free PMC article.
-
Replication Rapidly Recovers and Continues in the Presence of Hydroxyurea in Escherichia coli.J Bacteriol. 2018 Feb 23;200(6):e00713-17. doi: 10.1128/JB.00713-17. Print 2018 Mar 15. J Bacteriol. 2018. PMID: 29263100 Free PMC article.
-
Reactive oxygen species accelerate de novo acquisition of antibiotic resistance in E. coli.iScience. 2023 Oct 31;26(12):108373. doi: 10.1016/j.isci.2023.108373. eCollection 2023 Dec 15. iScience. 2023. PMID: 38025768 Free PMC article.
References
-
- Wallace SS. Biological consequences of free radical-damaged DNA bases. Free Radic Biol Med. 2002;33:1–14. - PubMed
-
- Ide H, Petrullo LA, Hatahet Z, Wallace SS. Processing of DNA base damage by DNA polymerases. Dihydrothymine and beta-ureidoisobutyric acid as models for instructive and noninstructive lesions. J Biol Chem. 1991;266:1469–1477. - PubMed
-
- Evans J, Maccabee M, Hatahet Z, Courcelle J, Bockrath R, Ide H, Wallace S. Thymine ring saturation and fragmentation products: lesion bypass, misinsertion and implications for mutagenesis. Mutat Res. 1993;299:147–156. - PubMed
-
- Efrati E, Tocco G, Eritja R, Wilson SH, Goodman MF. “Action-at-a-distance” mutagenesis. 8-oxo-7, 8-dihydro-2′-deoxyguanosine causes base substitution errors at neighboring template sites when copied by DNA polymerase beta. J Biol Chem. 1999;274:15920–15926. - PubMed
-
- Wallace SS. DNA damages processed by base excision repair: biological consequences. Int J Radiat Biol. 1994;66:579–589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
