

Prospero and Pax2 combinatorially control neural cell fate decisions by modulating Ras- and Notch-dependent signaling
- PMID: 21539742
- PMCID: PMC3123624
- DOI: 10.1186/1749-8104-6-20
Prospero and Pax2 combinatorially control neural cell fate decisions by modulating Ras- and Notch-dependent signaling
Abstract
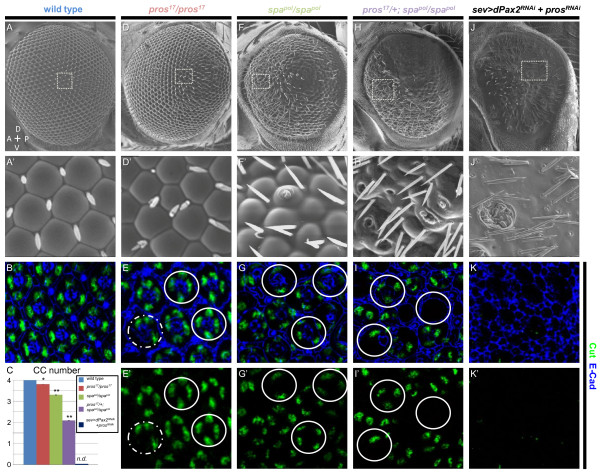
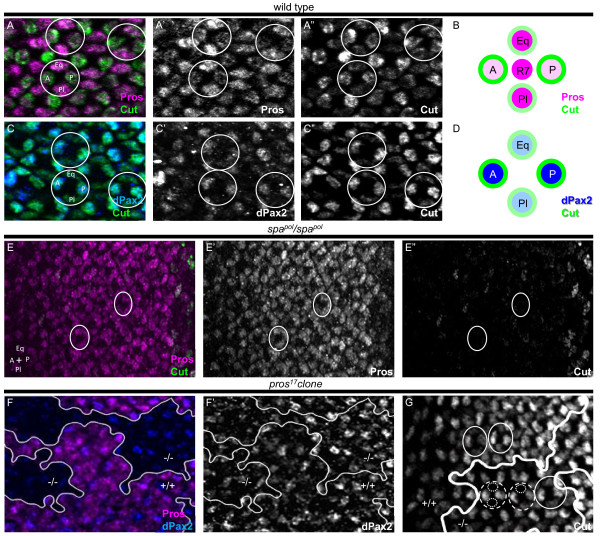
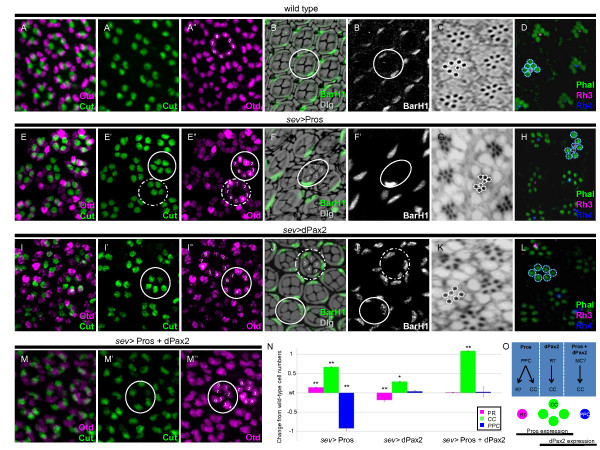
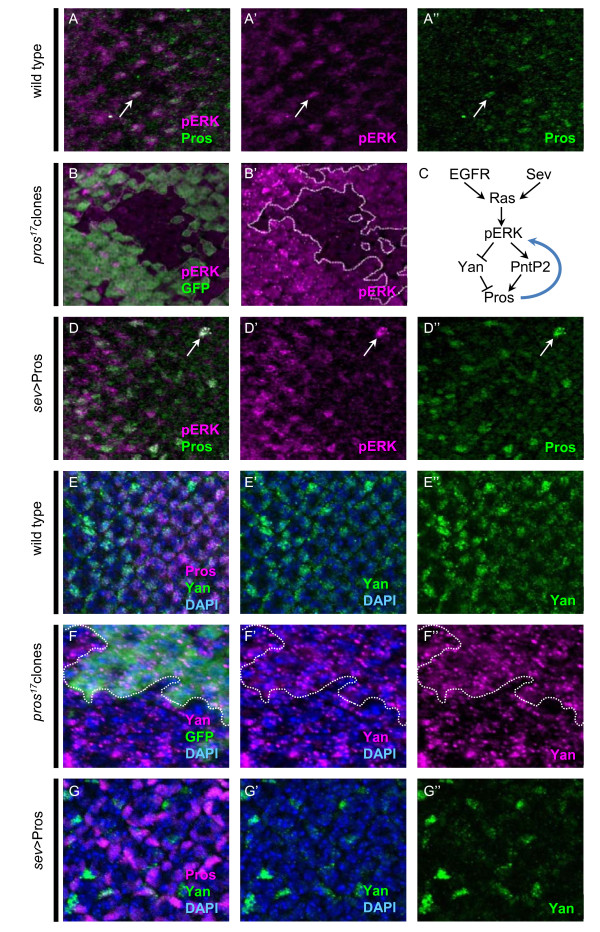
Background: The concept of an equivalence group, a cluster of cells with equal potential to adopt the same specific fate, has served as a useful paradigm to understand neural cell type specification. In the Drosophila eye, a set of five cells, called the 'R7 equivalence group', generates a single photoreceptor neuron and four lens-secreting epithelial cells. This choice between neuronal versus non-neuronal cell fates rests on differential requirements for, and cross-talk between, Notch/Delta- and Ras/mitogen-activated protein kinase (MAPK)-dependent signaling pathways. However, many questions remain unanswered related to how downstream events of these two signaling pathways mediate distinct cell fate decisions.
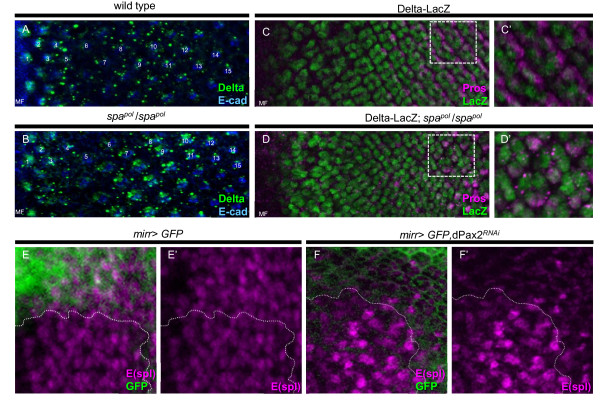
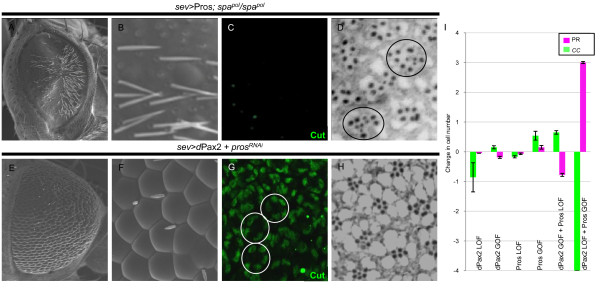
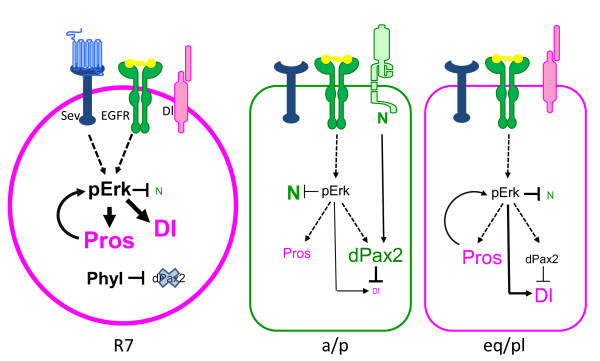
Results: Here, we demonstrate that two direct downstream targets of Ras and Notch signaling, the transcription factors Prospero and dPax2, are essential regulators of neuronal versus non-neuronal cell fate decisions in the R7 equivalence group. Prospero controls high activated MAPK levels required for neuronal fate, whereas dPax2 represses Delta expression to prevent neuronal fate. Importantly, activity from both factors is required for proper cell fate decisions to occur.
Conclusions: These data demonstrate that Ras and Notch signaling are integrated during cell fate decisions within the R7 equivalence group through the combinatorial and opposing activities of Pros and dPax2. Our study provides one of the first examples of how the differential expression and synergistic roles of two independent transcription factors determine cell fate within an equivalence group. Since the integration of Ras and Notch signaling is associated with many developmental and cancer models, these findings should provide new insights into how cell specificity is achieved by ubiquitously used signaling pathways in diverse biological contexts.
Figures
Similar articles
-
Determination of cell fates in the R7 equivalence group of the Drosophila eye by the concerted regulation of D-Pax2 and TTK88.Dev Biol. 2009 Jul 1;331(1):68-77. doi: 10.1016/j.ydbio.2009.04.026. Epub 2009 May 3. Dev Biol. 2009. PMID: 19406115
-
Parsimony and complexity: Cell fate assignment in the developing Drosophila eye.Fly (Austin). 2017 Jul 3;11(3):171-178. doi: 10.1080/19336934.2017.1291103. Epub 2017 Feb 6. Fly (Austin). 2017. PMID: 28165886 Free PMC article.
-
Three distinct roles for notch in Drosophila R7 photoreceptor specification.PLoS Biol. 2011 Aug;9(8):e1001132. doi: 10.1371/journal.pbio.1001132. Epub 2011 Aug 23. PLoS Biol. 2011. PMID: 21886484 Free PMC article.
-
Signal integration during development: insights from the Drosophila eye.Dev Dyn. 2004 Jan;229(1):162-75. doi: 10.1002/dvdy.10449. Dev Dyn. 2004. PMID: 14699588 Review.
-
Signal integration during development: mechanisms of EGFR and Notch pathway function and cross-talk.Crit Rev Biochem Mol Biol. 2006 Nov-Dec;41(6):339-85. doi: 10.1080/10409230600914344. Crit Rev Biochem Mol Biol. 2006. PMID: 17092823 Review.
Cited by
-
Two temporal functions of Glass: Ommatidium patterning and photoreceptor differentiation.Dev Biol. 2016 Jun 1;414(1):4-20. doi: 10.1016/j.ydbio.2016.04.012. Epub 2016 Apr 19. Dev Biol. 2016. PMID: 27105580 Free PMC article.
-
Glass promotes the differentiation of neuronal and non-neuronal cell types in the Drosophila eye.PLoS Genet. 2018 Jan 11;14(1):e1007173. doi: 10.1371/journal.pgen.1007173. eCollection 2018 Jan. PLoS Genet. 2018. PMID: 29324767 Free PMC article.
-
Eye Development in Sepia officinalis Embryo: What the Uncommon Gene Expression Profiles Tell Us about Eye Evolution.Front Physiol. 2017 Aug 24;8:613. doi: 10.3389/fphys.2017.00613. eCollection 2017. Front Physiol. 2017. PMID: 28883798 Free PMC article.
-
Hexagonal patterning of the Drosophila eye.Dev Biol. 2021 Oct;478:173-182. doi: 10.1016/j.ydbio.2021.07.004. Epub 2021 Jul 8. Dev Biol. 2021. PMID: 34245727 Free PMC article. Review.
-
Alterations of the CIB2 calcium- and integrin-binding protein cause Usher syndrome type 1J and nonsyndromic deafness DFNB48.Nat Genet. 2012 Nov;44(11):1265-71. doi: 10.1038/ng.2426. Epub 2012 Sep 30. Nat Genet. 2012. PMID: 23023331 Free PMC article.
References
-
- Frankfort BJ, Mardon G. R8 development in the Drosophila eye: a paradigm for neural selection and differentiation. Development. 2002;129:1295–1306. - PubMed
-
- Wolff T, Ready DF. Pattern Formation in the Drosophila Retina. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1993.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
