

A role for leukocyte-derived IL-1RA in DC homeostasis revealed by increased susceptibility of IL-1RA-deficient mice to cutaneous leishmaniasis
- PMID: 21525884
- PMCID: PMC6999703
- DOI: 10.1038/jid.2011.99
A role for leukocyte-derived IL-1RA in DC homeostasis revealed by increased susceptibility of IL-1RA-deficient mice to cutaneous leishmaniasis
Abstract
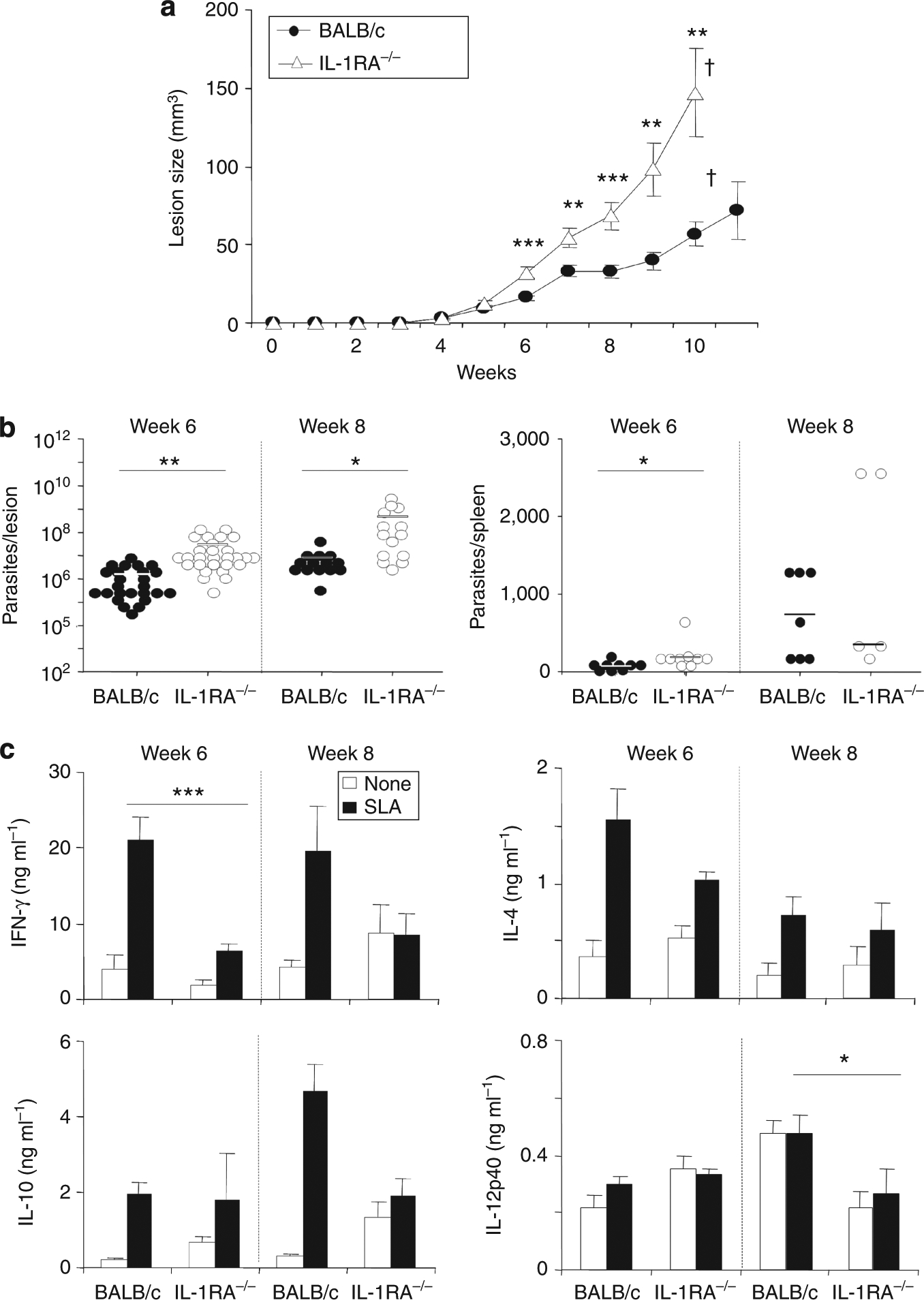
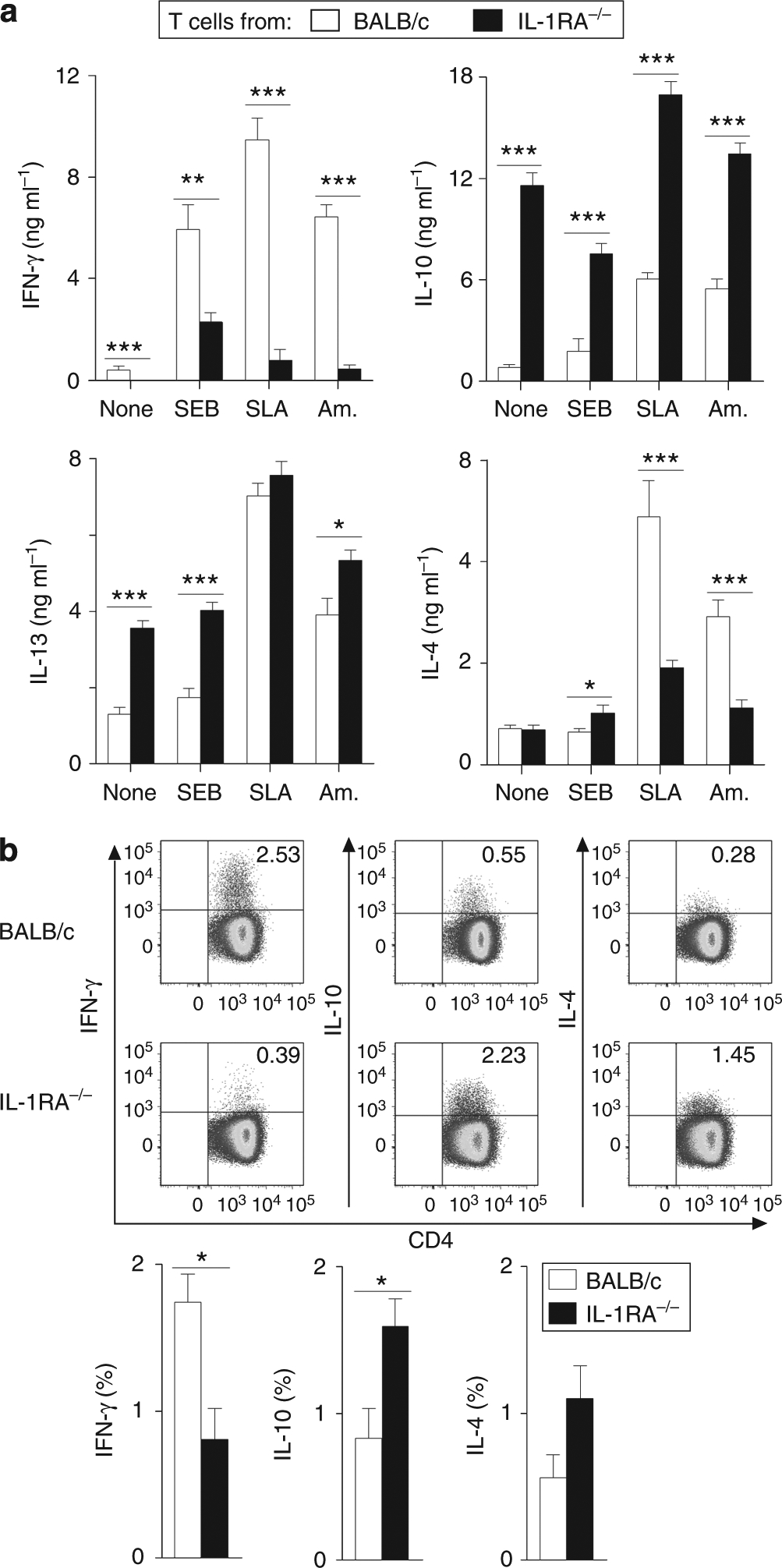
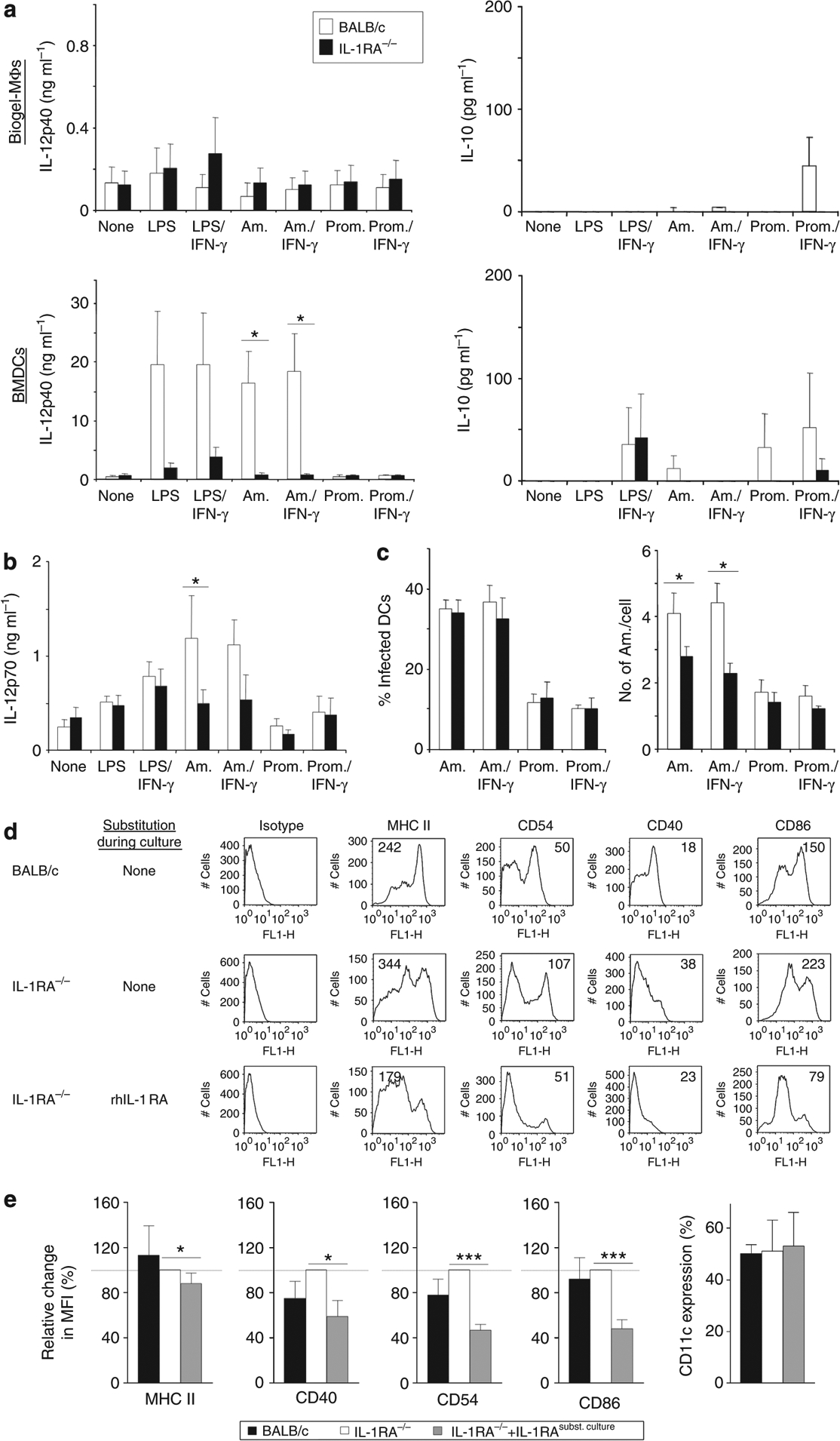
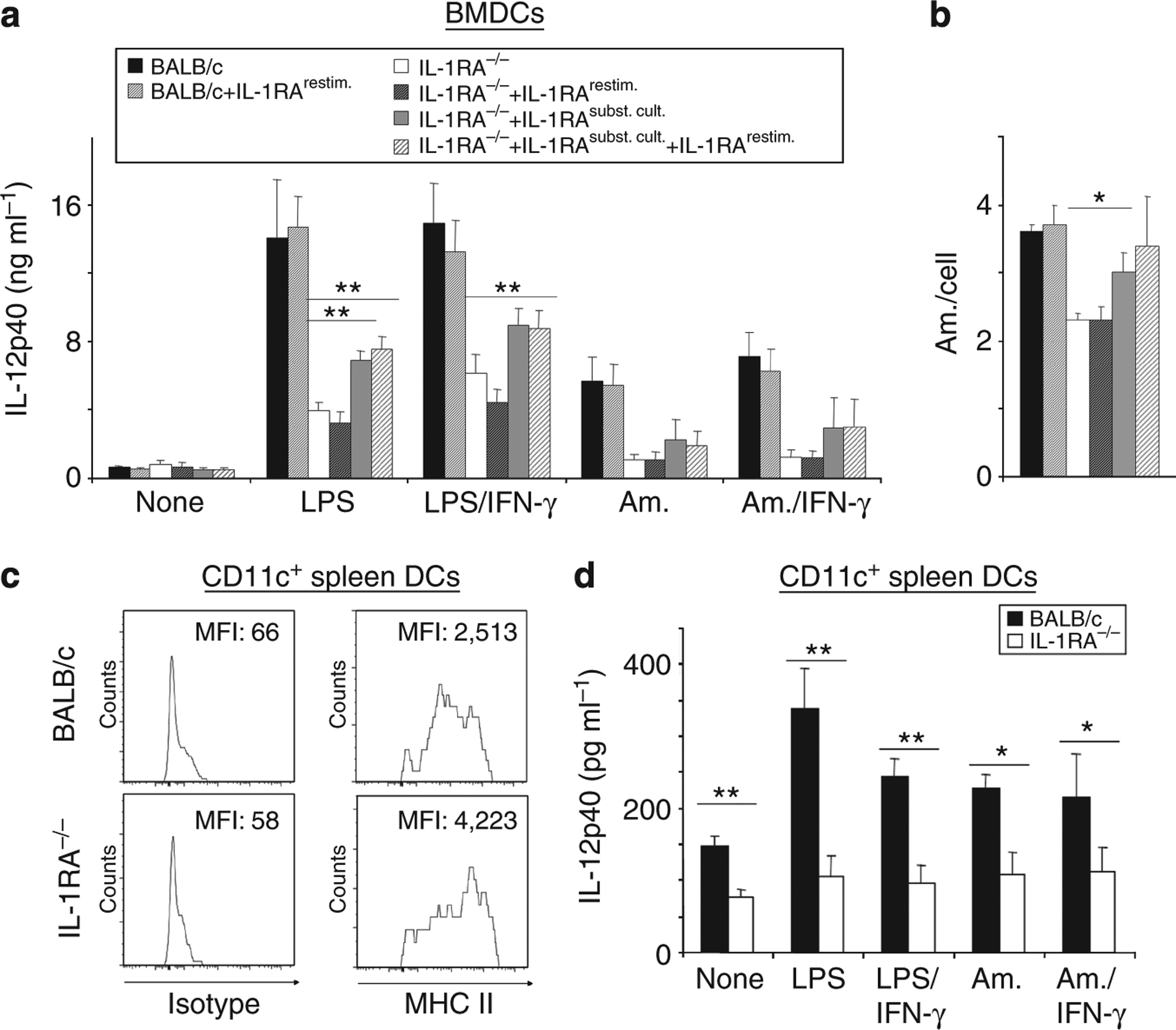
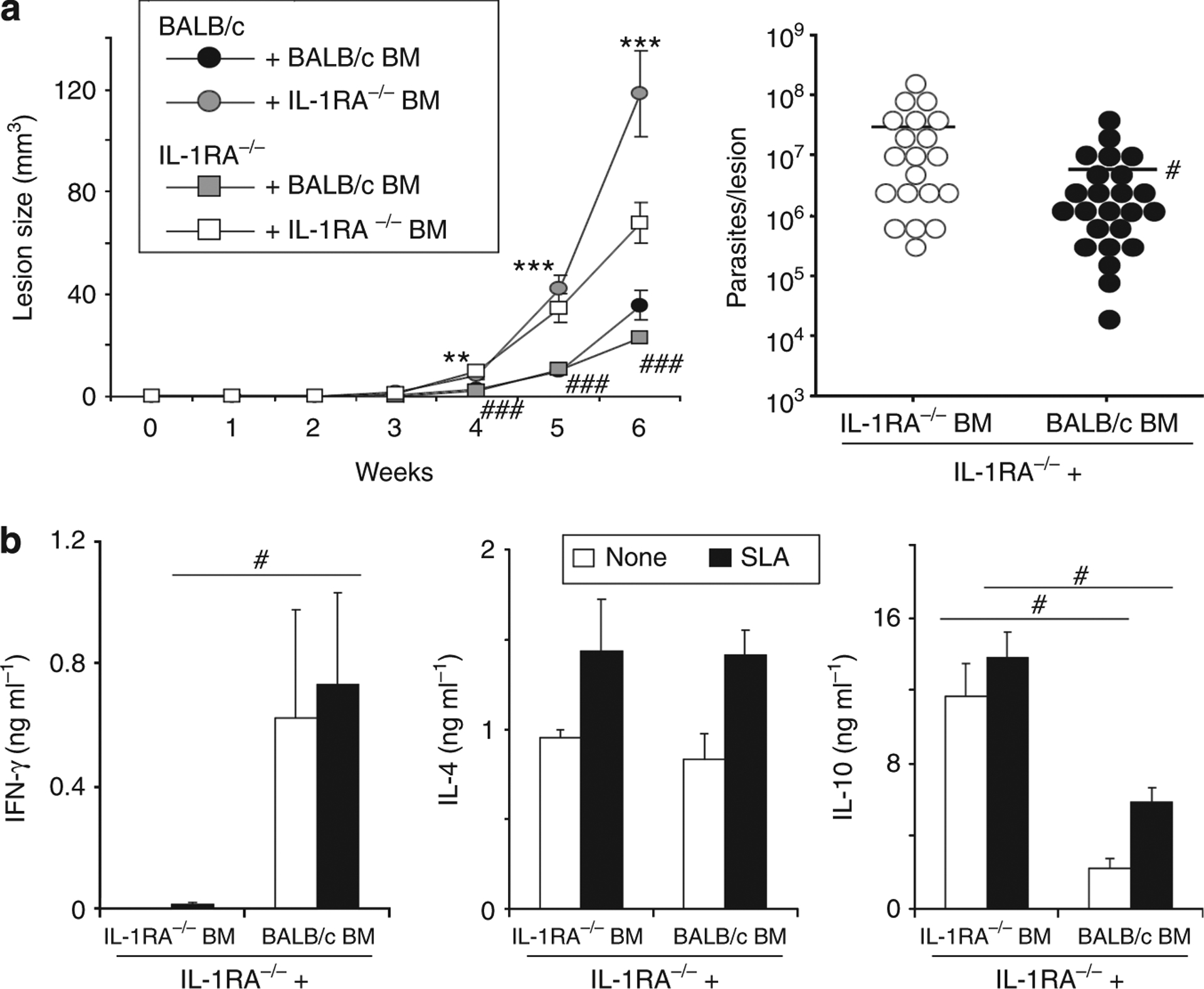
Dendritic cell (DC)-derived IL-1α/β plays a critical role in the induction of T helper type 1 (Th1)-dependent immunity against Leishmania. DCs from susceptible BALB/c mice produce less IL-1α/β when compared with resistant C57BL/6 mice, contributing to aberrant Th2 development and ultimate death of infected mice. We have extended our studies of the role of IL-1 in leishmaniasis using IL-1RA(-/-) BALB/c mice that are characterized by upregulated IL-1 receptor signaling. Unexpectedly, infection of IL-1RA(-/-) mice led to significantly worsened disease outcome with larger lesions, dramatically higher parasite burdens, and decreased IFN-γ production by antigen-specific T cells. We determined that IL-1RA(-/-) DCs were more mature already in the steady state, exhibited less phagocytotic capacity, and IL-12 production in response to various stimuli was impaired. Our data suggest that in addition to effects on Th education, IL-1α/β signaling also modulates DC homeostasis with increased signaling, leading to downmodulation of IL-12 synthesis and worsened disease outcome after infection with Leishmania major. Thus, the complex regulation of various members of the IL-1 cytokine family mediated through effects on both DCs and T cells critically contributes to disease outcome against this important human pathogen.
Conflict of interest statement
CONFLICT OF INTEREST
The authors state no conflict of interest.
Figures
Similar articles
-
Deletion of IL-4 receptor alpha on dendritic cells renders BALB/c mice hypersusceptible to Leishmania major infection.PLoS Pathog. 2013 Oct;9(10):e1003699. doi: 10.1371/journal.ppat.1003699. Epub 2013 Oct 24. PLoS Pathog. 2013. PMID: 24204259 Free PMC article.
-
Leishmania major-infected murine langerhans cell-like dendritic cells from susceptible mice release IL-12 after infection and vaccinate against experimental cutaneous Leishmaniasis.Eur J Immunol. 2000 Dec;30(12):3498-506. doi: 10.1002/1521-4141(2000012)30:12<3498::AID-IMMU3498>3.0.CO;2-6. Eur J Immunol. 2000. PMID: 11093169
-
Batf3-dependent CD103+ dendritic cells are major producers of IL-12 that drive local Th1 immunity against Leishmania major infection in mice.Eur J Immunol. 2015 Jan;45(1):119-29. doi: 10.1002/eji.201444651. Epub 2014 Nov 28. Eur J Immunol. 2015. PMID: 25312824 Free PMC article.
-
Anti-leishmania effector functions of CD4+ Th1 cells and early events instructing Th2 cell development and susceptibility to Leishmania major in BALB/c mice.Adv Exp Med Biol. 1998;452:53-60. doi: 10.1007/978-1-4615-5355-7_7. Adv Exp Med Biol. 1998. PMID: 9889959 Review.
-
Requirements for Th1-dependent immunity against infection with Leishmania major.Microbes Infect. 2004 Oct;6(12):1102-9. doi: 10.1016/j.micinf.2004.05.024. Microbes Infect. 2004. PMID: 15380780 Review.
Cited by
-
Variable gene expression and parasite load predict treatment outcome in cutaneous leishmaniasis.Sci Transl Med. 2019 Nov 20;11(519):eaax4204. doi: 10.1126/scitranslmed.aax4204. Sci Transl Med. 2019. PMID: 31748229 Free PMC article.
-
Host-Directed Therapies for Cutaneous Leishmaniasis.Front Immunol. 2021 Mar 26;12:660183. doi: 10.3389/fimmu.2021.660183. eCollection 2021. Front Immunol. 2021. PMID: 33841444 Free PMC article. Review.
-
Multiomic profiling of cutaneous leishmaniasis infections reveals microbiota-driven mechanisms underlying disease severity.Sci Transl Med. 2023 Oct 18;15(718):eadh1469. doi: 10.1126/scitranslmed.adh1469. Epub 2023 Oct 18. Sci Transl Med. 2023. PMID: 37851822 Free PMC article.
-
Immune responses against protozoan parasites: a focus on the emerging role of Nod-like receptors.Cell Mol Life Sci. 2016 Aug;73(16):3035-51. doi: 10.1007/s00018-016-2212-3. Epub 2016 Mar 31. Cell Mol Life Sci. 2016. PMID: 27032699 Free PMC article. Review.
-
Deletion of IL-4 receptor alpha on dendritic cells renders BALB/c mice hypersusceptible to Leishmania major infection.PLoS Pathog. 2013 Oct;9(10):e1003699. doi: 10.1371/journal.ppat.1003699. Epub 2013 Oct 24. PLoS Pathog. 2013. PMID: 24204259 Free PMC article.
References
-
- Belkaid Y, Butcher B, Sacks DL (1998) Analysis of cytokine production by inflammatory mouse macrophages at the single-cell level: selective impairment of IL-12 induction in Leishmania-infected cells. Eur J Immunol 28:1389–400 - PubMed
-
- Belkaid Y, Mendez S, Lira R et al. (2000) A natural model of Leishmania major infection reveals a prolonged “silent” phase of parasite amplification in the skin before the onset of lesion formation and immunity. J Immunol 165:969–77 - PubMed
-
- Belkaid Y, Von Stebut E, Mendez S et al. (2002) CD8+ T cells are required for primary immunity in C57BL/6 mice following low-dose, intradermal challenge with Leishmania major. J Immunol 168:3992–4000 - PubMed
-
- Berberich C, Ramirez-Pineda JR, Hambrecht C et al. (2003) Dendritic cell (DC)-based protection against an intracellular pathogen is dependent upon DC-derived IL-12 and can be induced by molecularly defined antigens. J Immunol 170:3171–9 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
