

Mechanisms controlling Th17 cytokine expression and host defense
- PMID: 21486905
- PMCID: PMC3133441
- DOI: 10.1189/jlb.0211099
Mechanisms controlling Th17 cytokine expression and host defense
Abstract
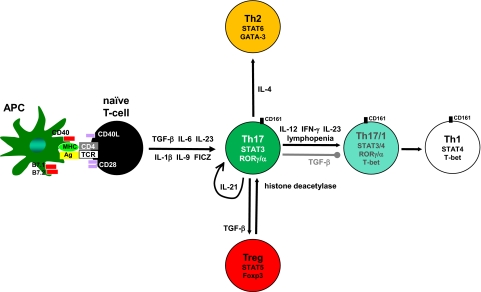
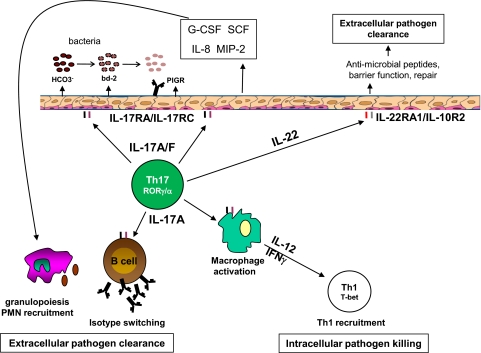
Th17 cells contribute to mucosal immunity by stimulating epithelial cells to induce antimicrobial peptides, granulopoiesis, neutrophil recruitment, and tissue repair. Recent studies have identified important roles for commensal microbiota and Ahr ligands in stabilizing Th17 gene expression in vivo, linking environmental cues to CD4 T cell polarization. Epigenetic changes that occur during the transition from naïve to effector Th17 cells increase the accessibility of il17a, il17f, and il22 loci to transcription factors. In addition, Th17 cells maintain the potential for expressing T-bet, Foxp3, or GATA-binding protein-3, explaining their plastic nature under various cytokine microenvironments. Although CD4 T cells are major sources of IL-17 and IL-22, innate cell populations, including γδ T cells, NK cells, and lymphoid tissue-inducer cells, are early sources of these cytokines during IL-23-driven responses. Epithelial cells and fibroblasts are important cellular targets for IL-17 in vivo; however, recent data suggest that macrophages and B cells are also stimulated directly by IL-17. Thus, Th17 cells interact with multiple populations to facilitate protection against intracellular and extracellular pathogens.
Figures
Similar articles
-
Protective role of Th17 cells in pulmonary infection.Vaccine. 2016 Mar 18;34(13):1504-1514. doi: 10.1016/j.vaccine.2016.02.021. Epub 2016 Feb 13. Vaccine. 2016. PMID: 26878294 Review.
-
Functional plasticity of Th17 cells: implications in gastrointestinal tract function.Int Rev Immunol. 2013 Oct-Dec;32(5-6):493-510. doi: 10.3109/08830185.2013.834899. Epub 2013 Sep 16. Int Rev Immunol. 2013. PMID: 24040751 Review.
-
Th17 cells and mucosal host defense.Semin Immunol. 2007 Dec;19(6):377-82. doi: 10.1016/j.smim.2007.10.009. Epub 2007 Nov 28. Semin Immunol. 2007. PMID: 18054248 Free PMC article. Review.
-
Cytokine signals through PI-3 kinase pathway modulate Th17 cytokine production by CCR6+ human memory T cells.J Exp Med. 2011 Aug 29;208(9):1875-87. doi: 10.1084/jem.20102516. Epub 2011 Aug 8. J Exp Med. 2011. PMID: 21825017 Free PMC article.
-
Microbial-induced Th17: superhero or supervillain?J Immunol. 2012 Oct 1;189(7):3285-91. doi: 10.4049/jimmunol.1201834. J Immunol. 2012. PMID: 22997231 Free PMC article. Review.
Cited by
-
Associations of genotypes and haplotypes of IL-17 with risk of gastric cancer in an eastern Chinese population.Oncotarget. 2016 Dec 13;7(50):82384-82395. doi: 10.18632/oncotarget.11616. Oncotarget. 2016. PMID: 27577072 Free PMC article.
-
Interleukin-22 mediates early host defense against Rhizomucor pusilluscan pathogens.PLoS One. 2013 Jun 17;8(6):e65065. doi: 10.1371/journal.pone.0065065. Print 2013. PLoS One. 2013. PMID: 23798999 Free PMC article.
-
Th1 and Th17 cells regulate innate immune responses and bacterial clearance during central nervous system infection.J Immunol. 2012 Feb 1;188(3):1360-70. doi: 10.4049/jimmunol.1101660. Epub 2011 Dec 21. J Immunol. 2012. PMID: 22190181 Free PMC article.
-
The role of IL-6 in host defence against infections: immunobiology and clinical implications.Nat Rev Rheumatol. 2017 Jul;13(7):399-409. doi: 10.1038/nrrheum.2017.83. Epub 2017 Jun 15. Nat Rev Rheumatol. 2017. PMID: 28615731 Review.
-
Leukemia inhibitory factor signaling is required for lung protection during pneumonia.J Immunol. 2012 Jun 15;188(12):6300-8. doi: 10.4049/jimmunol.1200256. Epub 2012 May 11. J Immunol. 2012. PMID: 22581855 Free PMC article.
References
-
- Hsu H. C., Yang P., Wang J., Wu Q., Myers R., Chen J., Yi J., Guentert T., Tousson A., Stanus A. L., et al. (2008) Interleukin 17-producing T helper cells and interleukin 17 orchestrate autoreactive germinal center development in autoimmune BXD2 mice. Nat. Immunol. 9, 166–175 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
