

Thermodynamic analysis of mutant lac repressors
- PMID: 21459098
- PMCID: PMC4225698
- DOI: 10.1016/j.jmb.2011.03.057
Thermodynamic analysis of mutant lac repressors
Abstract
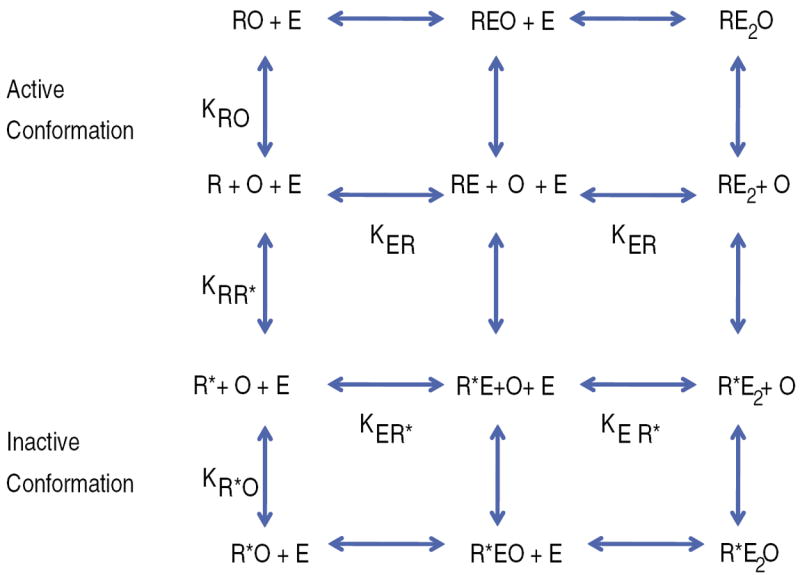
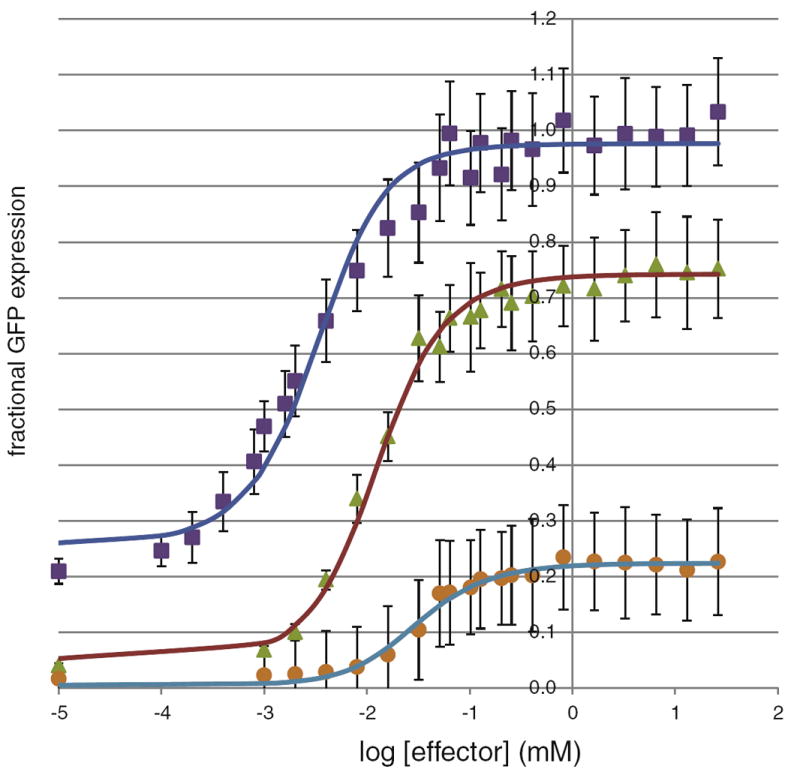
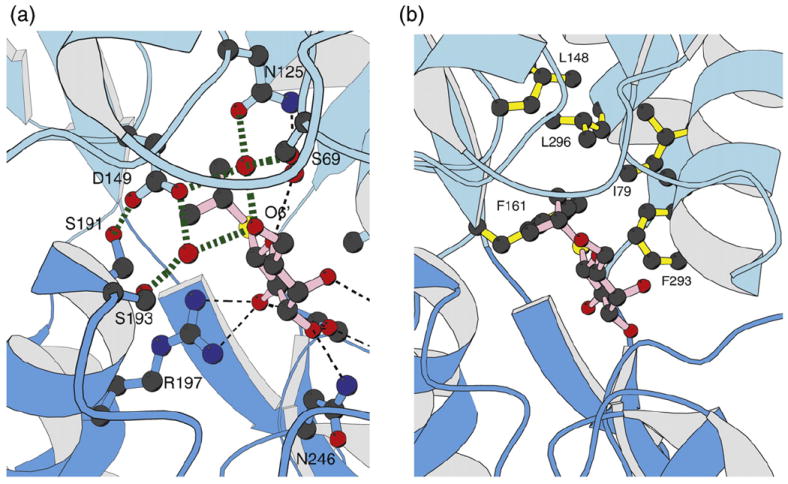
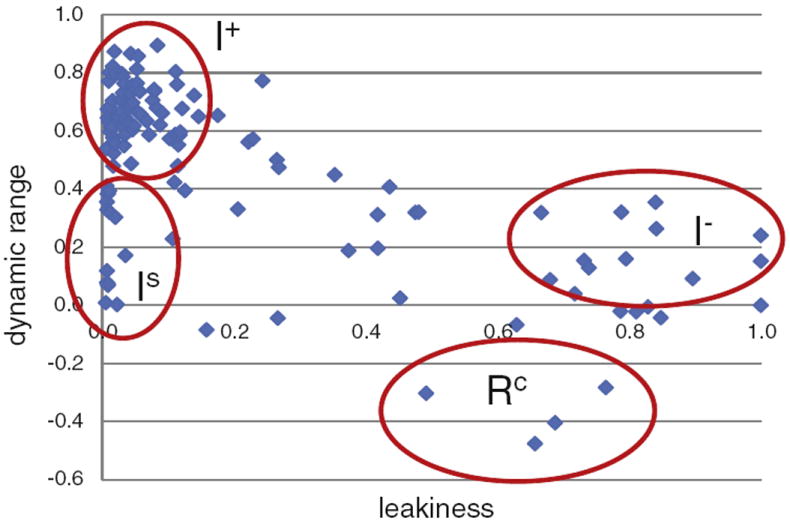
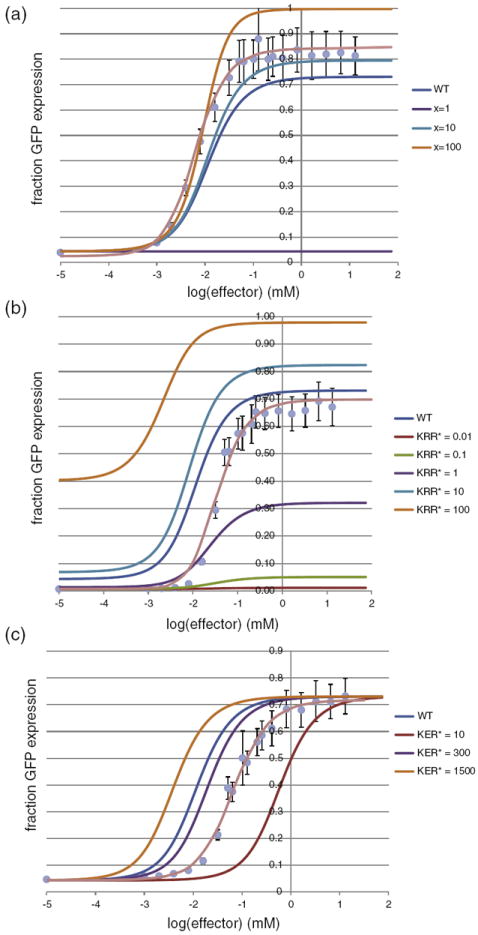
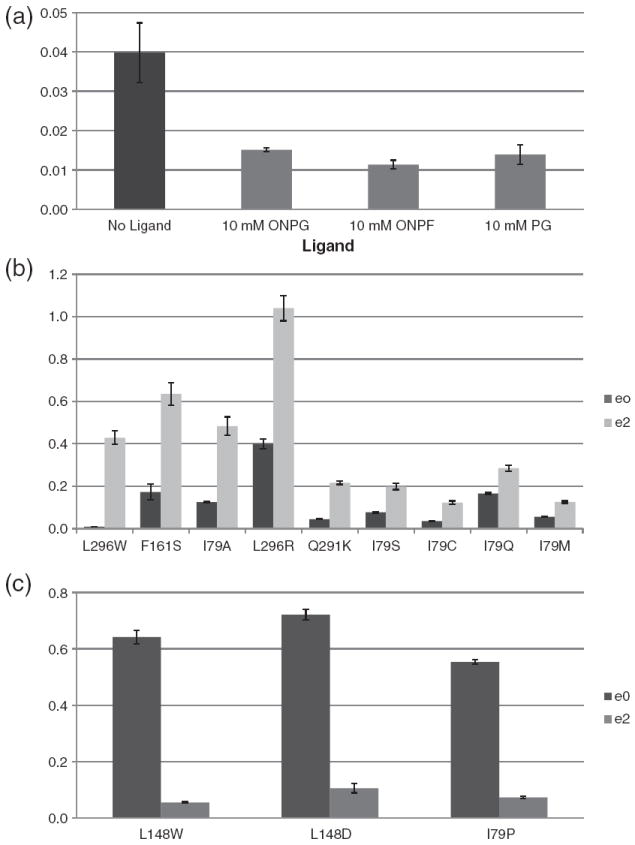
The lactose (lac) repressor is an allosteric protein that can respond to environmental changes. Mutations introduced into the DNA binding domain and the effector binding pocket affect the repressor's ability to respond to its environment. We have demonstrated how the observed phenotype is a consequence of altering the thermodynamic equilibrium constants. We discuss mutant repressors, which (1) show tighter repression; (2) induce with a previously noninducing species, orthonitrophenyl-β-D-galactoside; and (3) transform an inducible switch to one that is corepressed. The ability of point mutations to change multiple thermodynamic constants, and hence drastically alter the repressor's phenotype, shows how allosteric proteins can perform a wide array of similar yet distinct functions such as that exhibited in the Lac/Gal family of bacterial repressors.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
A single mutation in the core domain of the lac repressor reduces leakiness.Microb Cell Fact. 2013 Jul 8;12:67. doi: 10.1186/1475-2859-12-67. Microb Cell Fact. 2013. PMID: 23834731 Free PMC article.
-
Engineering alternate cooperative-communications in the lactose repressor protein scaffold.Protein Eng Des Sel. 2013 Jun;26(6):433-43. doi: 10.1093/protein/gzt013. Epub 2013 Apr 14. Protein Eng Des Sel. 2013. PMID: 23587523
-
Altering residues N125 and D149 impacts sugar effector binding and allosteric parameters in Escherichia coli lactose repressor.Biochemistry. 2011 Oct 25;50(42):9002-13. doi: 10.1021/bi200896t. Epub 2011 Sep 30. Biochemistry. 2011. PMID: 21928765 Free PMC article.
-
The lac repressor.C R Biol. 2005 Jun;328(6):521-48. doi: 10.1016/j.crvi.2005.04.004. C R Biol. 2005. PMID: 15950160 Review.
-
Lac repressor-operator complex.Curr Opin Struct Biol. 1997 Feb;7(1):76-85. doi: 10.1016/s0959-440x(97)80010-3. Curr Opin Struct Biol. 1997. PMID: 9032054 Review.
Cited by
-
The genotype-phenotype landscape of an allosteric protein.Mol Syst Biol. 2021 Mar;17(3):e10179. doi: 10.15252/msb.202010179. Mol Syst Biol. 2021. PMID: 33784029 Free PMC article.
-
Quantifying the regulatory role of individual transcription factors in Escherichia coli.Cell Rep. 2021 Nov 9;37(6):109952. doi: 10.1016/j.celrep.2021.109952. Cell Rep. 2021. PMID: 34758318 Free PMC article.
-
Predictive shifts in free energy couple mutations to their phenotypic consequences.Proc Natl Acad Sci U S A. 2019 Sep 10;116(37):18275-18284. doi: 10.1073/pnas.1907869116. Epub 2019 Aug 26. Proc Natl Acad Sci U S A. 2019. PMID: 31451655 Free PMC article.
-
A parametrized two-domain thermodynamic model explains diverse mutational effects on protein allostery.bioRxiv [Preprint]. 2024 Feb 14:2023.08.06.552196. doi: 10.1101/2023.08.06.552196. bioRxiv. 2024. Update in: Elife. 2024 Jun 05;12:RP92262. doi: 10.7554/eLife.92262 PMID: 37662419 Free PMC article. Updated. Preprint.
-
Tuning Transcriptional Regulation through Signaling: A Predictive Theory of Allosteric Induction.Cell Syst. 2018 Apr 25;6(4):456-469.e10. doi: 10.1016/j.cels.2018.02.004. Epub 2018 Mar 21. Cell Syst. 2018. PMID: 29574055 Free PMC article.
References
-
- Monod J, Changeux JP, Jacob F. Allosteric proteins and cellular control systems. J Mol Biol. 1963;6:306–329. - PubMed
-
- Monod J, Wyman J, Changeux JP. On the nature of allosteric transitions: a plausible model. J Mol Biol. 1965;12:88–118. - PubMed
-
- Jobe A, Bourgeois S. Lac repressor–operator interaction: VIII. Lactose is an anti-inducer of the lac operon. J Mol Biol. 1973;75:303–313. - PubMed
-
- Lewis M, Chang G, Horton NC, Kercher MA, Pace HC, Schumacher MA, et al. Crystal structure of the lactose operon repressor and its complexes with DNA and inducer. Science. 1996;271:1247–1254. see comment. - PubMed
-
- Boelens R, Scheek RM, van Boom JH, Kaptein R. Complex of lac repressor headpiece with a 14 base-pair lac operator fragment studied by twodimensional nuclear magnetic resonance. J Mol Biol. 1987;193:213–216. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
