

Citrullination of inhibitor of growth 4 (ING4) by peptidylarginine deminase 4 (PAD4) disrupts the interaction between ING4 and p53
- PMID: 21454715
- PMCID: PMC3089551
- DOI: 10.1074/jbc.M111.230961
Citrullination of inhibitor of growth 4 (ING4) by peptidylarginine deminase 4 (PAD4) disrupts the interaction between ING4 and p53
Abstract
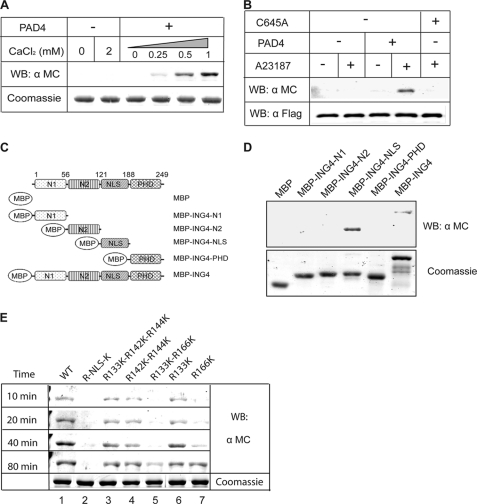
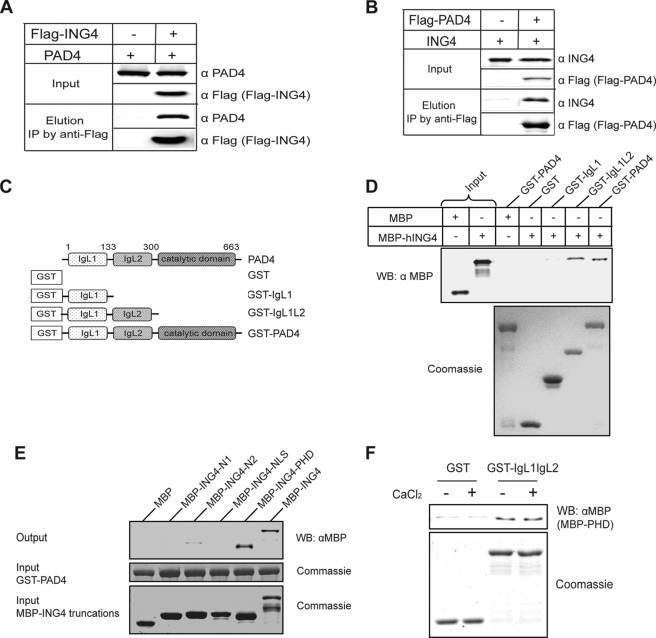
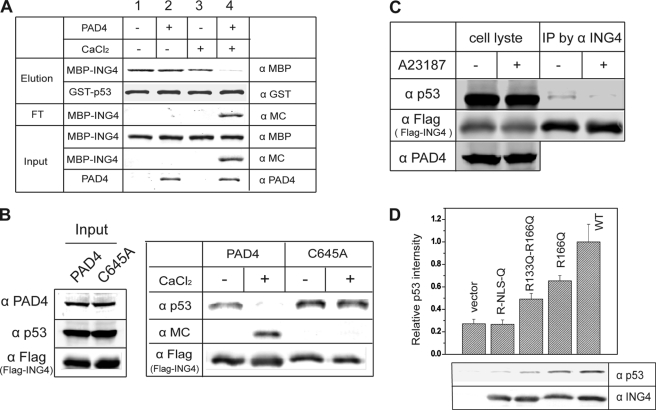
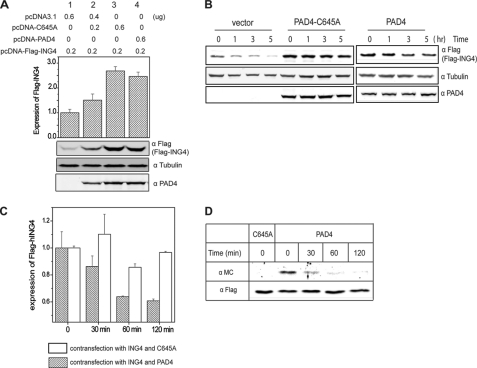
Gene expression is regulated by a number of interrelated posttranslational modifications of histones, including citrullination. For example, peptidylarginine deminase 4 (PAD4) converts peptidyl arginine to citrulline in histone H3 and can repress gene expression. However, regulation of gene expression through citrullination of non-histone proteins is less well defined. Herein, we identify a tumor suppressor protein, inhibitor of growth 4 (ING4), as a novel non-histone substrate of PAD4. ING4 is known to bind p53 via its nuclear localization signal (NLS) region and to enhance transcriptional activity of p53. We show that PAD4 preferentially citrullinates ING4 in the same NLS region and thereby disrupts the interaction between ING4 and p53. A citrulline-mimicking Arg-NLS-Gln ING4 mutant, which has all Arg residues in the NLS mutated to Gln, loses its affinity for p53, can no longer promote p53 acetylation, and results in repression of downstream p21 expression. In addition, we found that citrullination leads to increased susceptibility of ING4 to degradation, likely impacting p53-independent pathways as well. These findings elucidate an interaction between posttranslational citrullination, acetylation, and methylation and highlight an unusual mechanism whereby citrullination of a non-histone protein impacts gene regulation.
Figures


Similar articles
-
Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression.Oncogene. 2010 May 27;29(21):3153-62. doi: 10.1038/onc.2010.51. Epub 2010 Mar 1. Oncogene. 2010. PMID: 20190809 Free PMC article.
-
Regulation of p53 target gene expression by peptidylarginine deiminase 4.Mol Cell Biol. 2008 Aug;28(15):4745-58. doi: 10.1128/MCB.01747-07. Epub 2008 May 27. Mol Cell Biol. 2008. PMID: 18505818 Free PMC article.
-
Discovery of peptidylarginine deiminase-4 substrates by protein array: antagonistic citrullination and methylation of human ribosomal protein S2.Mol Biosyst. 2011 Jul;7(7):2286-95. doi: 10.1039/c1mb05089c. Epub 2011 May 16. Mol Biosyst. 2011. PMID: 21584310 Free PMC article.
-
Role of citrullination modification catalyzed by peptidylarginine deiminase 4 in gene transcriptional regulation.Acta Biochim Biophys Sin (Shanghai). 2017 Jul 1;49(7):567-572. doi: 10.1093/abbs/gmx042. Acta Biochim Biophys Sin (Shanghai). 2017. PMID: 28472221 Review.
-
Histone citrullination by protein arginine deiminase: is arginine methylation a green light or a roadblock?ACS Chem Biol. 2006 Aug 22;1(7):433-41. doi: 10.1021/cb6002306. ACS Chem Biol. 2006. PMID: 17168521 Review.
Cited by
-
Protein citrullination as a source of cancer neoantigens.J Immunother Cancer. 2021 Jun;9(6):e002549. doi: 10.1136/jitc-2021-002549. J Immunother Cancer. 2021. PMID: 34112737 Free PMC article.
-
PAD4 takes charge during neutrophil activation: Impact of PAD4 mediated NET formation on immune-mediated disease.J Thromb Haemost. 2021 Jul;19(7):1607-1617. doi: 10.1111/jth.15313. Epub 2021 May 12. J Thromb Haemost. 2021. PMID: 33773016 Free PMC article. Review.
-
Citrullination and the protein code: crosstalk between post-translational modifications in cancer.Philos Trans R Soc Lond B Biol Sci. 2023 Nov 20;378(1890):20220243. doi: 10.1098/rstb.2022.0243. Epub 2023 Oct 2. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37778382 Free PMC article. Review.
-
Inactivation of epidermal growth factor by Porphyromonas gingivalis as a potential mechanism for periodontal tissue damage.Infect Immun. 2013 Jan;81(1):55-64. doi: 10.1128/IAI.00830-12. Epub 2012 Oct 22. Infect Immun. 2013. PMID: 23090954 Free PMC article.
-
Peptidylarginine deiminase 2 plays a key role in osteogenesis by enhancing RUNX2 stability through citrullination.Cell Death Dis. 2023 Aug 30;14(8):576. doi: 10.1038/s41419-023-06101-7. Cell Death Dis. 2023. PMID: 37648716 Free PMC article.
References
-
- Tarcsa E., Marekov L. N., Mei G., Melino G., Lee S. C., Steinert P. M. (1996) J. Biol. Chem. 271, 30709–30716 - PubMed
-
- Pritzker L. B., Joshi S., Gowan J. J., Harauz G., Moscarello M. A. (2000) Biochemistry 39, 5374–5381 - PubMed
-
- Klose R. J., Zhang Y. (2007) Nat. Rev. Mol. Cell Biol. 8, 307–318 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
