

Review
doi: 10.1038/nrmicro2541.
Genome-virome interactions: examining the role of common viral infections in complex disease
Affiliations
- PMID: 21407242
- PMCID: PMC3678363
- DOI: 10.1038/nrmicro2541
Item in Clipboard
Review
Genome-virome interactions: examining the role of common viral infections in complex disease
Nat Rev Microbiol.
2011 Apr.
Abstract
New technologies have widened our view of 'complex diseases': those with both genetic and environmental risk factors. In this Review, we explore recent genetic and virological evidence implicating host-virus interactions in three diseases: type 1 diabetes, inflammatory bowel disease and asthma. The viruses implicated in these diseases cause mucosal infections that affect most of the population but are asymptomatic or mild in many hosts. These findings place a new emphasis on common viral infections as important environmental factors in the pathogenesis of complex diseases, and they compel the field to pursue a better understanding of host interactions with the human virome.
Figures
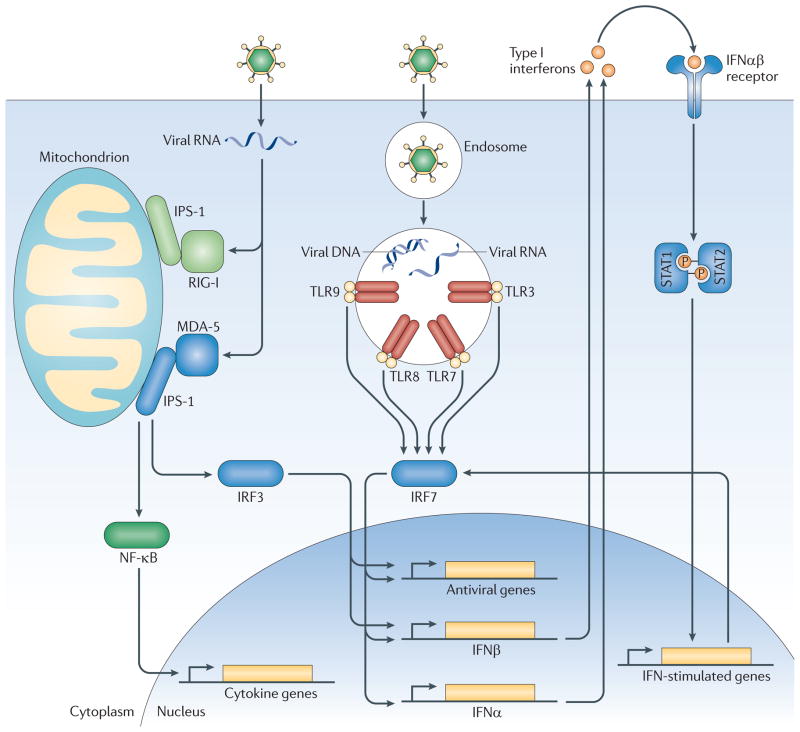
Viruses are recognized in general by two separate pathways. Intrinsic recognition occurs through detection of viral nucleic acids by cytosolic RLRs and other nucleic acid sensors in the infected cells. MDA-5/IFI1H is a cytosolic RLR that can recognize Picornavirus genomes. Activated MDA-5 activates the transcription factors IRF-3 and NFκB. In contrast, extrinsic recognition of virus occurs through Toll-like receptors (TLR) 3, 7, 8 and 9, which can recognize viral DNA and RNA within endosomes and activate transcription factors IRF-7 and NF-κb. IRF-3 and IRF-7 function in homo- or heterodimers to initiate transcription of Type I interferons and other anti-viral genes, as shown. Via NF-κB, both pathways induce the expression of pro-inflammatory cytokines. Secreted Type I interferons bind to the IFNαβR on the cell surface, which signals via STAT1 and STAT2 to induce expression of ~300 interferon-stimulated genes (ISGs) including MDA-5 and IRF-7. Type 1 diabetes is linked to genetic polymorphisms in MDA-5/IFI1H, IRF-7, and an IRF-7 driven network of 305 genes (likely the depicted network of IFN response genes.) Interestingly, T1D is predicted to correlate with robust responses through these pathways. For MDA-5, polymorphisms that result greater expression correlate with disease, whereas rare alleles that result in loss of or reduced function are protective. Similarly, T1D-associated polymorphisms in the genomic locus regulating the IRF-7 driven network predict increased expression of this network in T1D.
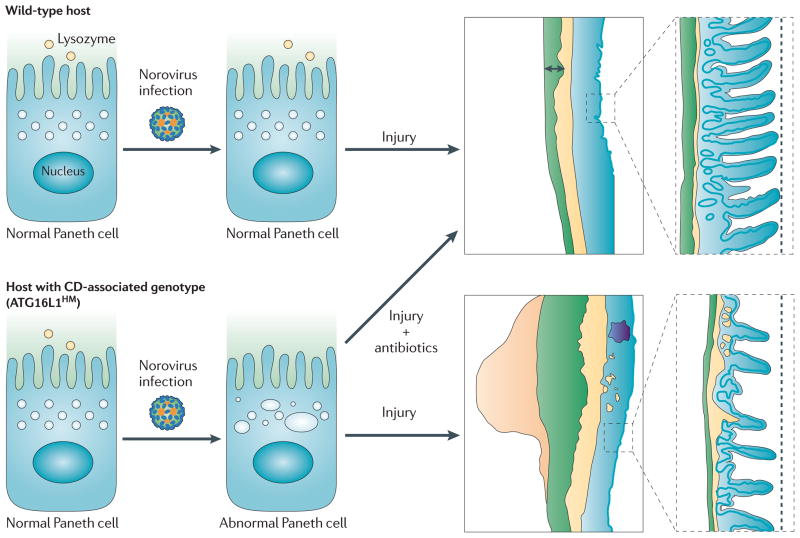
A mouse model of Crohn’s disease (CD) requires both a genetic predisposition and a viral infection for the expression of the disease phenotype. In this model, Virgin and colleagues compared intestinal physiology in wild-type hosts and in hosts with reduced expression of a Crohn’s disease-associated autophagy gene, ATG16L1. Wild-type hosts display the predicted intestinal histology during recovery from an intestinal injury. Similarly, hosts deficient in ATG16L1 have normal-appearing intestinal Paneth cells and the usual response to intestinal injury if norovirus infection is absent. However, if ATG16L1-mutant hosts are infected with a persistent strain of norovirus, they develop abnormal Paneth cells and have an abnormal response to intestinal injury with histopathology resembling Crohn’s disease. As illustrated, the injured colon in these mice colon displays lymphoid aggregates, abnormal thickening of the muscularis propria, and submucosa inflammation. In parallel, the intestinal injury agent induced abnormal histology in the ileum of mice with the predisposing gene + norovirus infection, but not in wild-type norovirus infected mice. Of note, CD-like intestinal histopathology does not develop if genetically-predisposed, virus infected mice are treated with antibiotics, indicating that the intestinal microbiota is also an important environmental factor for disease expression.
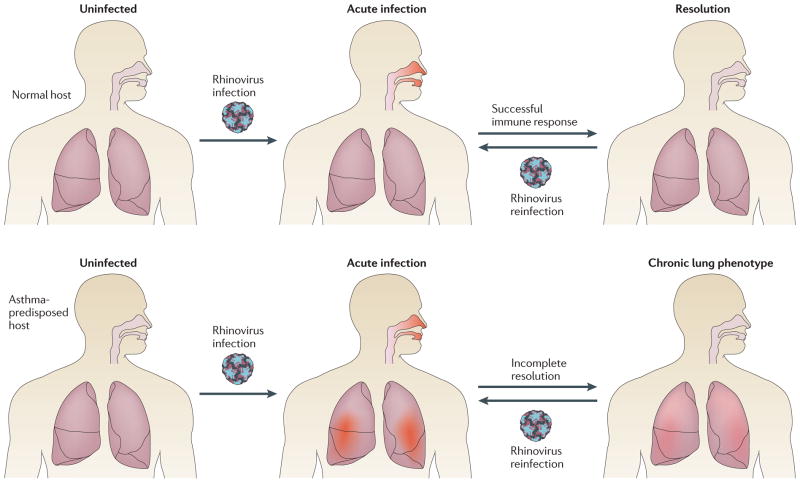
PCR-based studies have revealed a high incidence and prevalence of rhinovirus respiratory infections. At the same time, rhinovirus has been identified as a major precipitant of asthma exacerbations, and growing evidence indicates that asthmatic subjects develop more severe symptoms upon rhinovirus infection than healthy controls. This diagram illustrates how genetic polymorphisms affecting the host rhinovirus response could contribute to asthma pathogenesis. In this simplified scenario, all hosts experience frequent, repeated rhinovirus infections throughout their lifetimes (at least once every few months as children.) These infections may be asymptomatic, result in mild symptoms of the common cold, or result in upper and lower airway inflammation with wheezing and respiratory distress. In normal hosts, rhinovirus infections affect only the upper airway, and are mild or asymptomatic. Each infection fully resolves and leaves airways intact. In asthma-predisposed hosts, genetic polymorphisms in the antiviral immune response result in more prolonged and severe rhinovirus infections, with both upper and lower airway inflammation. In addition to acute episodes, repeated more severe infections lead to chronic inflammation and airway remodeling over time. This example illustrates how a common and frequent viral infection, in the setting of a detrimental host response, could contribute to the pathogenesis of asthma.
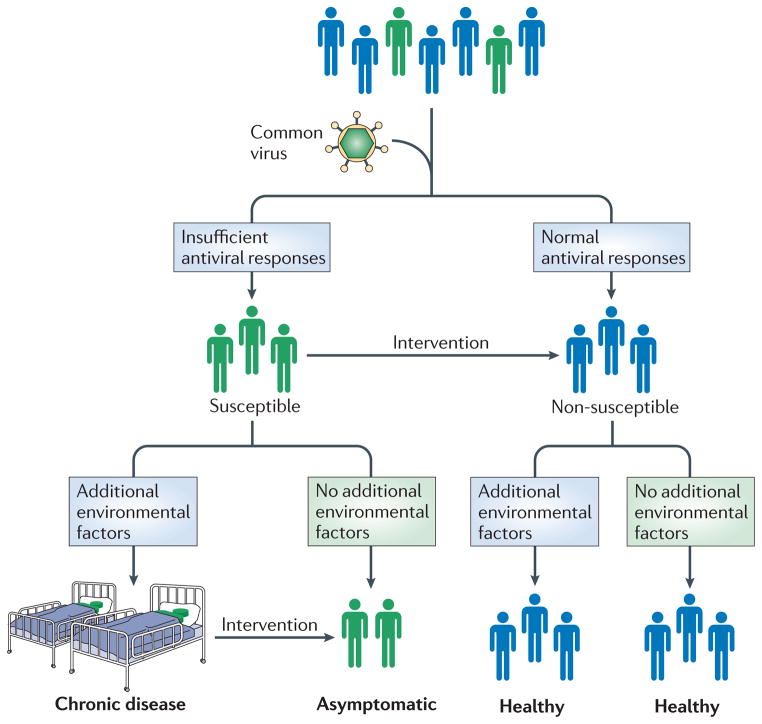
Converging evidence suggests that variations in the host antiviral response are important features of complex disease pathogenesis, in the context of a constant-albeit-changing virome of common infections. In this model, all hosts are exposed to certain highly prevalent viruses. Those with a non-susceptible antiviral response do not get the disease. Those with an altered antiviral response get the disease, but only if certain other key environmental factors are present. Avoiding these key environmental factors offers one avenue for circumventing complex diseases. This model also suggests a new mode for intervention in complex disease pathogenesis. If we understood the mechanisms whereby the altered antiviral response led to disease, we could potentially design interventions to shift this response to that of a non-predisposed host. Such interventions might include vaccines, well-timed virus exposures, or other immune modulating therapeutics. This model suggests that to tackle complex diseases, it is critical to learn more about the human virome and factors that influence host-virome interactions.
Similar articles
-
The Role of Viruses in the Pathogenesis of Immune-Mediated Gastro-Intestinal Diseases.Int J Mol Sci. 2024 Jul 30;25(15):8301. doi: 10.3390/ijms25158301. Int J Mol Sci. 2024. PMID: 39125870 Free PMC article. Review.
-
The virome in host health and disease.Immunity. 2015 May 19;42(5):805-13. doi: 10.1016/j.immuni.2015.05.003. Immunity. 2015. PMID: 25992857 Free PMC article. Review.
-
The science of the host-virus network.Nat Microbiol. 2021 Dec;6(12):1483-1492. doi: 10.1038/s41564-021-00999-5. Epub 2021 Nov 24. Nat Microbiol. 2021. PMID: 34819645 Review.
-
Viral manipulation of the host immune response.Curr Opin Immunol. 2015 Oct;36:54-60. doi: 10.1016/j.coi.2015.06.012. Epub 2015 Jul 10. Curr Opin Immunol. 2015. PMID: 26177523 Free PMC article. Review.
-
Chemical Methods for Probing Virus-Host Proteomic Interactions.ACS Infect Dis. 2016 Nov 11;2(11):773-786. doi: 10.1021/acsinfecdis.6b00084. Epub 2016 Jun 29. ACS Infect Dis. 2016. PMID: 27933785 Review.
Cited by
-
Interactions between Bacteriophage, Bacteria, and the Mammalian Immune System.Viruses. 2018 Dec 25;11(1):10. doi: 10.3390/v11010010. Viruses. 2018. PMID: 30585199 Free PMC article. Review.
-
Beyond Cytomegalovirus and Epstein-Barr Virus: a Review of Viruses Composing the Blood Virome of Solid Organ Transplant and Hematopoietic Stem Cell Transplant Recipients.Clin Microbiol Rev. 2020 Aug 26;33(4):e00027-20. doi: 10.1128/CMR.00027-20. Print 2020 Sep 16. Clin Microbiol Rev. 2020. PMID: 32847820 Free PMC article. Review.
-
Beyond research: a primer for considerations on using viral metagenomics in the field and clinic.Front Microbiol. 2015 Mar 25;6:224. doi: 10.3389/fmicb.2015.00224. eCollection 2015. Front Microbiol. 2015. PMID: 25859244 Free PMC article.
-
A Role for the Intestinal Microbiota and Virome in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS)?J Clin Med. 2016 Jun 6;5(6):55. doi: 10.3390/jcm5060055. J Clin Med. 2016. PMID: 27275835 Free PMC article. Review.
-
Invasions of Host-Associated Microbiome Networks.Adv Ecol Res. 2017;57:201-281. doi: 10.1016/bs.aecr.2016.11.002. Adv Ecol Res. 2017. PMID: 39404686 Free PMC article.
References
-
- Mullis KB, Faloona FA. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Meth Enzymol. 1987;155:335–350. - PubMed
-
- Mackay IM, Mackay JF, Nissen MD, Sloots TP. In: Real-time PCR in Microbiology: From Diagnosis to Characterisation. Mackay IM, editor. Caister Academic Press; Norfolk, U. K: 2007. pp. 1–40.
-
- Monto AS, Cavallaro JJ. The Tecumseh study of respiratory illness. II Patterns of occurrence of infection with respiratory pathogens, 1965–1969. Am J Epidemiol. 1971;94:280–289. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
