

Review
doi: 10.1083/jcb.201010058.
Principles of chromosomal organization: lessons from yeast
Affiliations
- PMID: 21383075
- PMCID: PMC3051815
- DOI: 10.1083/jcb.201010058
Item in Clipboard
Review
Principles of chromosomal organization: lessons from yeast
J Cell Biol.
.
Abstract
The spatial organization of genes and chromosomes plays an important role in the regulation of several DNA processes. However, the principles and forces underlying this nonrandom organization are mostly unknown. Despite its small dimension, and thanks to new imaging and biochemical techniques, studies of the budding yeast nucleus have led to significant insights into chromosome arrangement and dynamics. The dynamic organization of the yeast genome during interphase argues for both the physical properties of the chromatin fiber and specific molecular interactions as drivers of nuclear order.
Figures

General configuration of yeast chromosomes in interphase yeast. (top) Subnuclear territories occupied by three loci and the SPB, obtained using the methodology and data described previously (Berger et al., 2008; Therizols et al., 2010). Thousands of nuclei were detected in 3D microscopy images and computationally oriented along a central axis (broken line in the bottom panel) defined by the nuclear center and the nucleolus center of mass (X, bottom), allowing the determination of both the radial distance of loci relative to the nuclear center and an elevation angle above the central axis. Each color represents a locus or the SPB; dark and light shades indicate high and low probabilities, respectively. Green, rDNA; blue, the SPB; red, subtelomere of the short (85 kb) chromosome arm 9R; cyan, subtelomere of the long (440 kb) chromosome arm 11L. Note how the rDNA, the site of nucleolar protein assembly, occupies a pole of the nucleus opposite the microtubule organization center (SPB). Bar, 1 µm. (bottom) Sketch of a Rabl-like chromosome configuration hypothesized on the basis of observed subnuclear positions (Therizols et al., 2010). Enlargements depict possible chromosome arrangements at smaller scales. The rightmost enlargement shows an array of nucleosomes in a loose chromatin fiber (Dekker, 2008), with a segment of nucleosome-free DNA looping out. The bottom panel is adapted from Therizols et al. (2010).
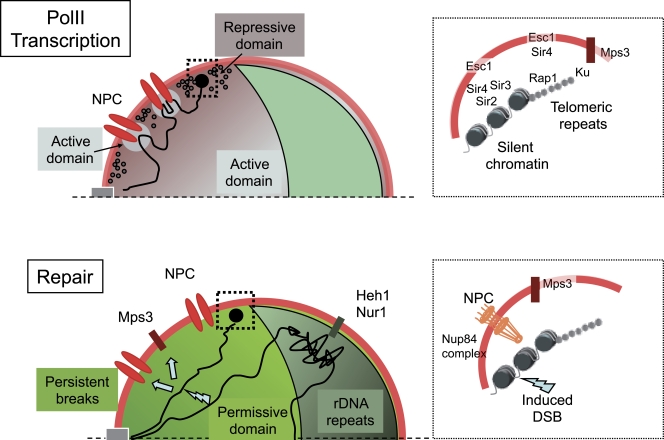
Functional compartmentalization of the yeast nucleus. (top) PolII transcription domains. The nucleolus, where polI and polIII transcription occur, is shown in green. The nucleoplasm is partitioned between polII transcriptionally repressive domains (dark gray), where silencing proteins (black circles) are found, and domains permissive for transcription (light gray), in the nuclear interior and in the proximity of NPCs (red). Chromatin loops (black line) accompany the activation of inducible genes. Large black dot, chromosome end. The inset on the right shows molecular determinants of transcriptional silencing acting at telomeric repeats and the upstream subtelomere. (bottom) DNA repair domains. Permanent breaks induced either by HO or I-SceI endonucleases (lightning bolt symbol) are displaced (arrows) to the nuclear periphery through the action of the Nup84 subcomplex or the Mps3 protein (Therizols et al., 2006; Nagai et al., 2008; Oza et al., 2009). This displacement requires Mec1 and modification of the histone H2AZ. At the NPC, modification of repair proteins by SUMO ubiquitin ligases modifying enzymes Slx5 and Slx8 or the desumoylase Ulp1 might be important to ensure efficient repair (not depicted; Palancade et al., 2007; Nagai et al., 2008). In the nucleolus, recombination between rDNA repeats is prevented by their association with Heh1 and Nur1, proteins of the inner nuclear membrane (Mekhail et al., 2008). The inset on the right shows a permanent break induced at subtelomeres displaced toward the NPC or Mps3.

Nuclear foci can occur in absence of interactions. Simulated microscopy images of fluorescently tagged telomeres (green) in a yeast nucleus (left) and quantification of the distribution of simulated foci number (right). (left, top) Telomeres are confined to a portion of the nuclear periphery. (left, bottom) telomeres are randomly distributed in the nucleoplasm. In each simulation, 32 telomeres were randomly positioned independently of each other within a subvolume of a 1-µm-radius spherical nucleus. In the top row, this subvolume represents the “nuclear periphery” and is a spherical shell of inner and outer radii of 0.8 µm and 1 µm, respectively. In the bottom row, this subvolume consists of the entire nuclear sphere with the exception of a region representing the nucleolus. The 3D orientation of the nucleus was chosen randomly for each panel. Red, nucleoplasm. To account for limited resolution, the point spread function was defined according to typical microscopy parameters, with full width at half maximum of ∼0.2 µm and 0.6 µm along the lateral and axial directions, respectively. The simulated images were corrupted by a mixture of Poisson and additive Gaussian noise. Each panel shows maximum intensity projections along the axial direction of a 3D image stack. Histograms of the number of foci were obtained from 2,000 independent simulations each (counts are on the y axis). Two telomeres located at a distance <0.3 µm were considered to be part of the same focus. Some foci contained only one telomere. The number of foci increases when telomeres are allowed to explore most of the nuclear volume. The histograms indicate substantially more distinct foci than are visible in the images, partly because the projection images merge spatially separated foci and because the signal of single isolated telomeres is difficult to detect visually.
Similar articles
-
Visualizing chromatin dynamics in interphase nuclei.Science. 2002 May 24;296(5572):1412-6. doi: 10.1126/science.1067703. Science. 2002. PMID: 12029120 Review.
-
Capturing chromosome conformation.Science. 2002 Feb 15;295(5558):1306-11. doi: 10.1126/science.1067799. Science. 2002. PMID: 11847345
-
Yeast nuclei display prominent centromere clustering that is reduced in nondividing cells and in meiotic prophase.J Cell Biol. 1998 Apr 6;141(1):21-9. doi: 10.1083/jcb.141.1.21. J Cell Biol. 1998. PMID: 9531545 Free PMC article.
-
Chromosome dynamics in the yeast interphase nucleus.Science. 2001 Dec 7;294(5549):2181-6. doi: 10.1126/science.1065366. Science. 2001. PMID: 11739961
-
The Yeast Genomes in Three Dimensions: Mechanisms and Functions.Annu Rev Genet. 2017 Nov 27;51:23-44. doi: 10.1146/annurev-genet-120116-023438. Epub 2017 Aug 30. Annu Rev Genet. 2017. PMID: 28853923 Review.
Cited by
-
High-Resolution Microscopy to Learn the Nuclear Organization of the Living Yeast Cells.Stem Cells Int. 2021 Aug 27;2021:9951114. doi: 10.1155/2021/9951114. eCollection 2021. Stem Cells Int. 2021. Retraction in: Stem Cells Int. 2024 Jan 24;2024:9872132. doi: 10.1155/2024/9872132. PMID: 34497652 Free PMC article. Retracted.
-
From dynamic chromatin architecture to DNA damage repair and back.Nucleus. 2018 Jan 1;9(1):161-170. doi: 10.1080/19491034.2017.1419847. Nucleus. 2018. PMID: 29271297 Free PMC article. Review.
-
The genome in space and time: does form always follow function? How does the spatial and temporal organization of a eukaryotic genome reflect and influence its functions?Bioessays. 2012 Sep;34(9):800-10. doi: 10.1002/bies.201200034. Epub 2012 Jul 6. Bioessays. 2012. PMID: 22777837 Free PMC article.
-
A decade of 3C technologies: insights into nuclear organization.Genes Dev. 2012 Jan 1;26(1):11-24. doi: 10.1101/gad.179804.111. Genes Dev. 2012. PMID: 22215806 Free PMC article. Review.
-
Destabilization of chromosome structure by histone H3 lysine 27 methylation.PLoS Genet. 2019 Apr 22;15(4):e1008093. doi: 10.1371/journal.pgen.1008093. eCollection 2019 Apr. PLoS Genet. 2019. PMID: 31009462 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
