

Zebrafish embryo screen for mycobacterial genes involved in the initiation of granuloma formation reveals a newly identified ESX-1 component
- PMID: 21372049
- PMCID: PMC3124061
- DOI: 10.1242/dmm.006676
Zebrafish embryo screen for mycobacterial genes involved in the initiation of granuloma formation reveals a newly identified ESX-1 component
Abstract
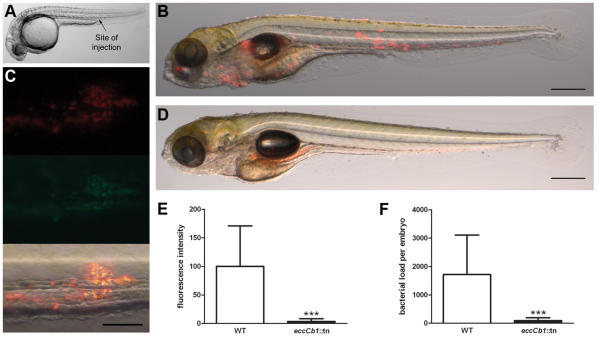
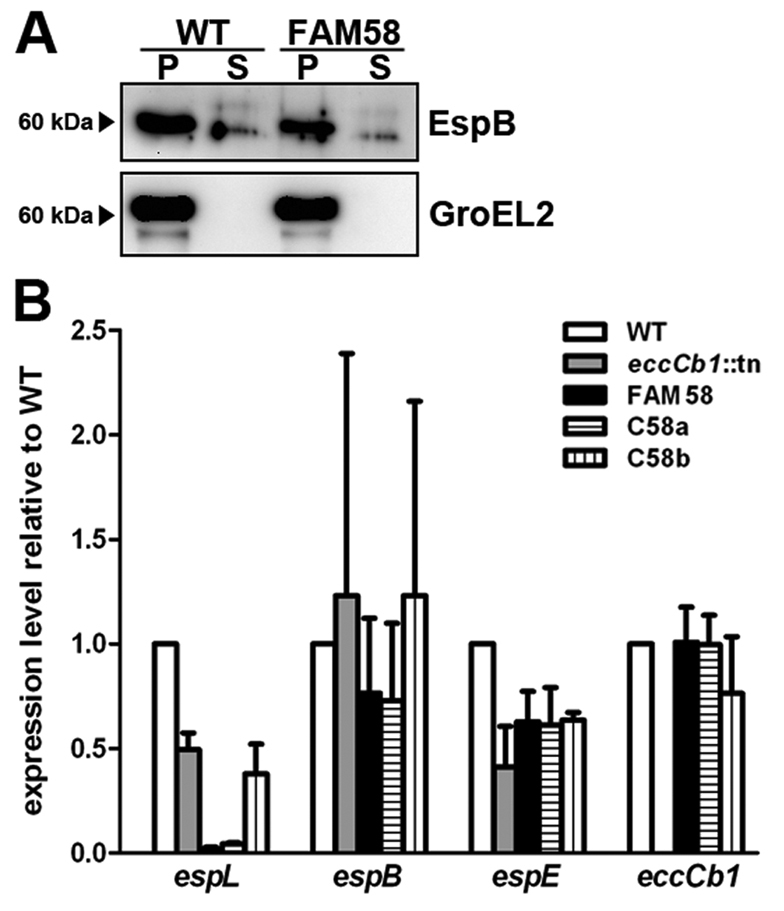
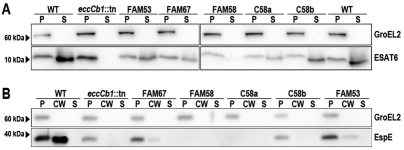
The hallmark of tuberculosis (TB) is the formation of granulomas, which are clusters of infected macrophages surrounded by additional macrophages, neutrophils and lymphocytes. Although it has long been thought that granulomas are beneficial for the host, there is evidence that mycobacteria also promote the formation of these structures. In this study, we aimed to identify new mycobacterial factors involved in the initial stages of granuloma formation. We exploited the zebrafish embryo Mycobacterium marinum infection model to study initiation of granuloma formation and developed an in vivo screen to select for random M. marinum mutants that were unable to induce granuloma formation efficiently. Upon screening 200 mutants, three mutants repeatedly initiated reduced granuloma formation. One of the mutants was found to be defective in the espL gene, which is located in the ESX-1 cluster. The ESX-1 cluster is disrupted in the Mycobacterium bovis BCG vaccine strain and encodes a specialized secretion system known to be important for granuloma formation and virulence. Although espL has not been implicated in protein secretion before, we observed a strong effect on the secretion of the ESX-1 substrates ESAT-6 and EspE. We conclude that our zebrafish embryo M. marinum screen is a useful tool to identify mycobacterial genes involved in the initial stages of granuloma formation and that we have identified a new component of the ESX-1 secretion system. We are confident that our approach will contribute to the knowledge of mycobacterial virulence and could be helpful for the development of new TB vaccines.
Figures



Similar articles
-
A mycobacterial virulence gene cluster extending RD1 is required for cytolysis, bacterial spreading and ESAT-6 secretion.Mol Microbiol. 2004 Sep;53(6):1677-93. doi: 10.1111/j.1365-2958.2004.04261.x. Mol Microbiol. 2004. PMID: 15341647
-
ESX-5-deficient Mycobacterium marinum is hypervirulent in adult zebrafish.Cell Microbiol. 2012 May;14(5):728-39. doi: 10.1111/j.1462-5822.2012.01755.x. Epub 2012 Feb 15. Cell Microbiol. 2012. PMID: 22256857
-
EspH is a hypervirulence factor for Mycobacterium marinum and essential for the secretion of the ESX-1 substrates EspE and EspF.PLoS Pathog. 2018 Aug 13;14(8):e1007247. doi: 10.1371/journal.ppat.1007247. eCollection 2018 Aug. PLoS Pathog. 2018. PMID: 30102741 Free PMC article.
-
New insights into the pathogenesis of tuberculosis revealed by Mycobacterium marinum: the zebrafish model from the systems biology perspective.Crit Rev Eukaryot Gene Expr. 2011;21(4):337-45. doi: 10.1615/critreveukargeneexpr.v21.i4.40. Crit Rev Eukaryot Gene Expr. 2011. PMID: 22181703 Review.
-
A fresh look at mycobacterial pathogenicity with the zebrafish host model.Mol Microbiol. 2022 Mar;117(3):661-669. doi: 10.1111/mmi.14838. Epub 2021 Nov 7. Mol Microbiol. 2022. PMID: 34714579 Free PMC article. Review.
Cited by
-
The CXCR3-CXCL11 signaling axis mediates macrophage recruitment and dissemination of mycobacterial infection.Dis Model Mech. 2015 Mar;8(3):253-69. doi: 10.1242/dmm.017756. Epub 2015 Jan 8. Dis Model Mech. 2015. PMID: 25573892 Free PMC article.
-
Deletion of a dehydratase important for intracellular growth and cording renders rough Mycobacterium abscessus avirulent.Proc Natl Acad Sci U S A. 2016 Jul 19;113(29):E4228-37. doi: 10.1073/pnas.1605477113. Epub 2016 Jul 6. Proc Natl Acad Sci U S A. 2016. PMID: 27385830 Free PMC article.
-
Mycobacteria employ two different mechanisms to cross the blood-brain barrier.Cell Microbiol. 2018 Sep;20(9):e12858. doi: 10.1111/cmi.12858. Epub 2018 May 30. Cell Microbiol. 2018. PMID: 29749044 Free PMC article.
-
Correlation of phenotypic profiles using targeted proteomics identifies mycobacterial esx-1 substrates.J Proteome Res. 2014 Nov 7;13(11):5151-64. doi: 10.1021/pr500484w. Epub 2014 Aug 25. J Proteome Res. 2014. PMID: 25106450 Free PMC article.
-
Infection and RNA-seq analysis of a zebrafish tlr2 mutant shows a broad function of this toll-like receptor in transcriptional and metabolic control and defense to Mycobacterium marinum infection.BMC Genomics. 2019 Nov 20;20(1):878. doi: 10.1186/s12864-019-6265-1. BMC Genomics. 2019. PMID: 31747871 Free PMC article.
References
-
- Abdallah A. M., Verboom T., Hannes F., Safi M., Strong M., Eisenberg D., Musters R. J., Vandenbroucke-Grauls C. M., Appelmelk B. J., Luirink J., et al. (2006). A specific secretion system mediates PPE41 transport in pathogenic mycobacteria. Mol. Microbiol. 62, 667–679 - PubMed
-
- Abdallah A. M., Verboom T., Weerdenburg E. M., Gey van Pittius N. C., Mahasha P. W., Jimenez C., Parra M., Cadieux N., Brennan M. J., Appelmelk B. J., et al. (2009). PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol. Microbiol. 73, 329–340 - PubMed
-
- Besra G. S. (1998). Preparation of cell-wall fractions from mycobacteria. In Methods in Molecular Biology: Mycobacteria Protocols (ed. Parish T., Stoker N. G.), pp. 91–107 Totowa, NJ: Humana Press - PubMed
-
- Bishai W. (2000). Lipid lunch for persistent pathogen. Nature 406, 683–685 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
