

Overexpression of BCL2 enhances survival of human embryonic stem cells during stress and obviates the requirement for serum factors
- PMID: 21300885
- PMCID: PMC3044421
- DOI: 10.1073/pnas.1019047108
Overexpression of BCL2 enhances survival of human embryonic stem cells during stress and obviates the requirement for serum factors
Abstract
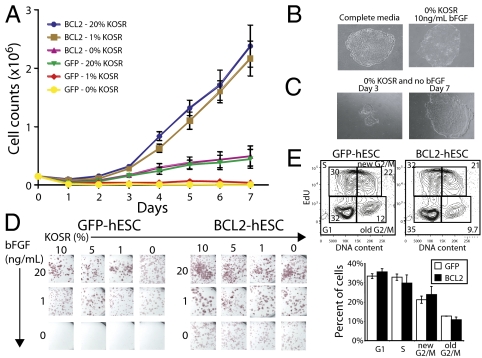
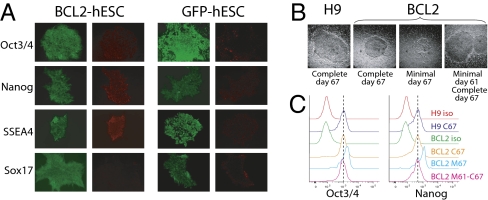
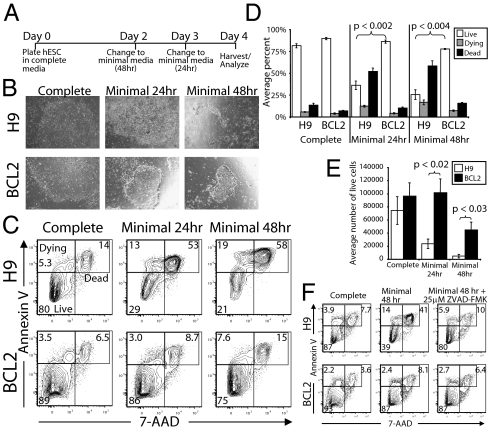
The promise of pluripotent stem cells as a research and therapeutic tool is partly undermined by the technical challenges of generating and maintaining these cells in culture. Human embryonic stem cells (hESCs) are exquisitely sensitive to culture conditions, and require constant signaling by growth factors and cell-cell and cell-matrix interactions to prevent apoptosis, senescence, and differentiation. Previous work from our laboratory demonstrated that overexpression of the prosurvival gene BCL2 in mouse embryonic stem cells overrode the requirement of serum factors and feeder cells to maintain mESCs in culture. To determine whether this prosurvival gene could similarly protect hESCs, we generated hESC lines that constitutively or inducibly express BCL2. We find that BCL2 overexpression significantly decreases dissociation-induced apoptosis, resulting in enhanced colony formation from sorted single cells, and enhanced embryoid body formation. In addition, BCL2-hESCs exhibit normal growth in the absence of serum, but require basic fibroblast growth factor to remain undifferentiated. Furthermore, they maintain their pluripotency markers, form teratomas in vivo, and differentiate into all three germ layers. Our data suggest that the BCL2 signaling pathway plays an important role in inhibiting hESC apoptosis, such that its overexpression in hESCs offers both a survival benefit in conditions of stress by resisting apoptosis and obviates the requirement for serum or a feeder layer for maintenance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Enhanced survival of pluripotent stem cells under stressful conditions.Cell Cycle. 2011 Aug 15;10(16):2610-1. doi: 10.4161/cc.10.16.16527. Epub 2011 Aug 15. Cell Cycle. 2011. PMID: 21791974 No abstract available.
Similar articles
-
Maintenance of human embryonic stem cells in media conditioned by human mesenchymal stem cells obviates the requirement of exogenous basic fibroblast growth factor supplementation.Tissue Eng Part C Methods. 2012 May;18(5):387-96. doi: 10.1089/ten.TEC.2011.0546. Epub 2012 Jan 26. Tissue Eng Part C Methods. 2012. PMID: 22136131
-
Undifferentiated propagation of the human embryonic stem cell lines, H1 and HSF6, on human placenta-derived feeder cells without basic fibroblast growth factor supplementation.Stem Cells Dev. 2010 Nov;19(11):1713-22. doi: 10.1089/scd.2010.0014. Epub 2010 Sep 9. Stem Cells Dev. 2010. PMID: 20201681
-
In vitro neural differentiation of human embryonic stem cells using a low-density mouse embryonic fibroblast feeder protocol.Methods Mol Biol. 2010;584:71-95. doi: 10.1007/978-1-60761-369-5_4. Methods Mol Biol. 2010. PMID: 19907972
-
A review of gene expression profiling of human embryonic stem cell lines and their differentiated progeny.Curr Stem Cell Res Ther. 2009 May;4(2):98-106. doi: 10.2174/157488809788167409. Curr Stem Cell Res Ther. 2009. PMID: 19442194 Review.
-
Homologous recombination in human embryonic stem cells: a tool for advancing cell therapy and understanding and treating human disease.Clin Transl Sci. 2011 Aug;4(4):298-305. doi: 10.1111/j.1752-8062.2011.00281.x. Clin Transl Sci. 2011. PMID: 21884519 Free PMC article. Review.
Cited by
-
Chimpanzee and pig-tailed macaque iPSCs: Improved culture and generation of primate cross-species embryos.Cell Rep. 2022 Aug 30;40(9):111264. doi: 10.1016/j.celrep.2022.111264. Cell Rep. 2022. PMID: 36044843 Free PMC article.
-
Integrated loss- and gain-of-function screens define a core network governing human embryonic stem cell behavior.Genes Dev. 2021 Nov 1;35(21-22):1527-1547. doi: 10.1101/gad.349048.121. Epub 2021 Oct 28. Genes Dev. 2021. PMID: 34711655 Free PMC article.
-
Apoptotic susceptibility to DNA damage of pluripotent stem cells facilitates pharmacologic purging of teratoma risk.Stem Cells Transl Med. 2012 Oct;1(10):709-18. doi: 10.5966/sctm.2012-0066. Epub 2012 Sep 27. Stem Cells Transl Med. 2012. PMID: 23197662 Free PMC article.
-
Methyl-CpG-binding domain 3 (Mbd3) is an important regulator for apoptosis in mouse embryonic stem cells.Am J Transl Res. 2020 Dec 15;12(12):8147-8161. eCollection 2020. Am J Transl Res. 2020. PMID: 33437388 Free PMC article.
-
AKT/GSK3β signaling pathway is critically involved in human pluripotent stem cell survival.Sci Rep. 2016 Oct 20;6:35660. doi: 10.1038/srep35660. Sci Rep. 2016. PMID: 27762303 Free PMC article.
References
-
- Hasegawa K, Fujioka T, Nakamura Y, Nakatsuji N, Suemori H. A method for the selection of human embryonic stem cell sublines with high replating efficiency after single-cell dissociation. Stem Cells. 2006;24:2649–2660. - PubMed
-
- Pyle AD, Lock LF, Donovan PJ. Neurotrophins mediate human embryonic stem cell survival. Nat Biotechnol. 2006;24:344–350. - PubMed
-
- Skottman H, Narkilahti S, Hovatta O. Challenges and approaches to the culture of pluripotent human embryonic stem cells. Regen Med. 2007;2:265–273. - PubMed
-
- Chin AC, Padmanabhan J, Oh SK, Choo AB. Defined and serum-free media support undifferentiated human embryonic stem cell growth. Stem Cells Dev. 2010;19:753–761. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
