

Ultrasensitivity in the Regulation of Cdc25C by Cdk1
- PMID: 21292159
- PMCID: PMC3060667
- DOI: 10.1016/j.molcel.2011.01.012
Ultrasensitivity in the Regulation of Cdc25C by Cdk1
Abstract
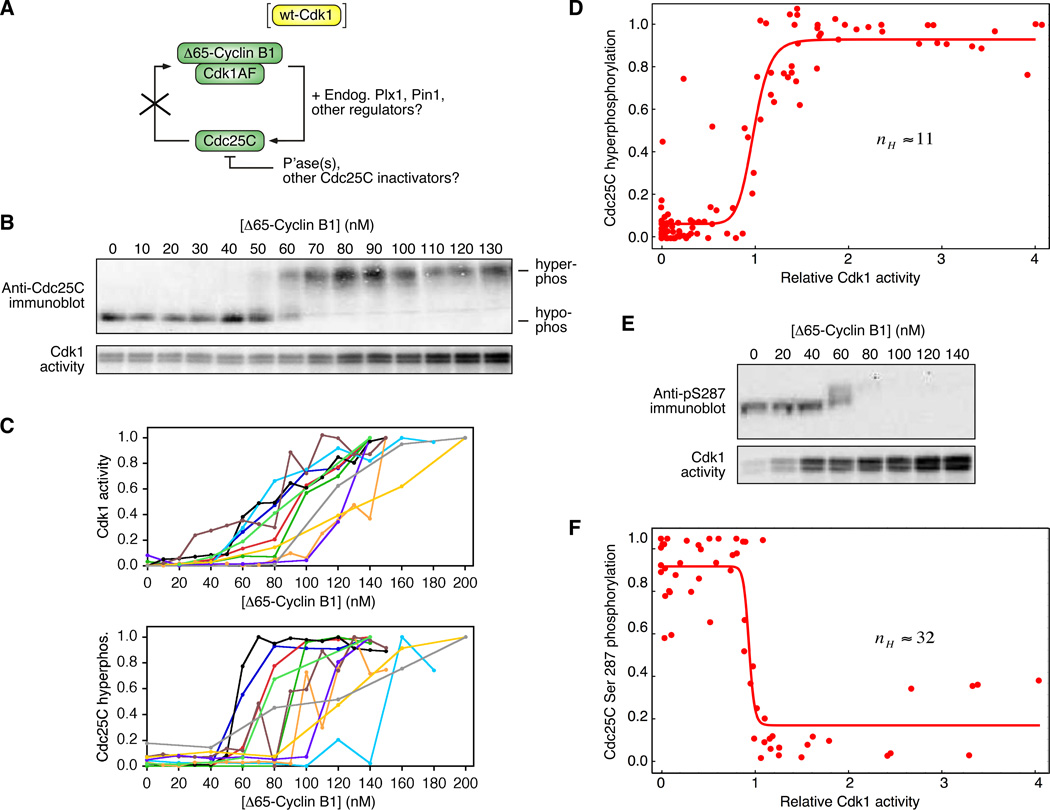
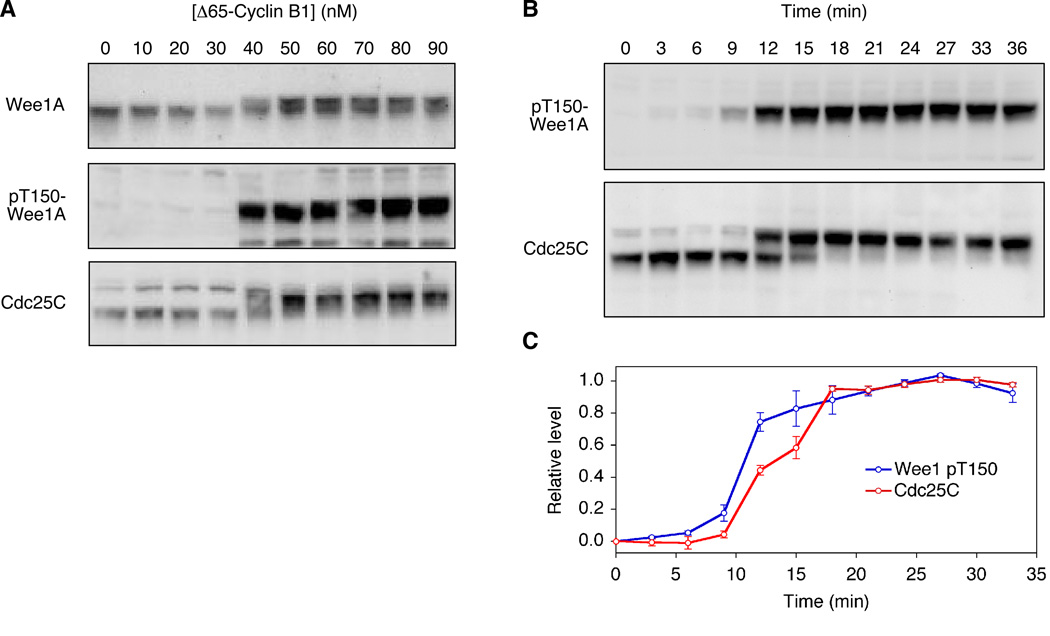
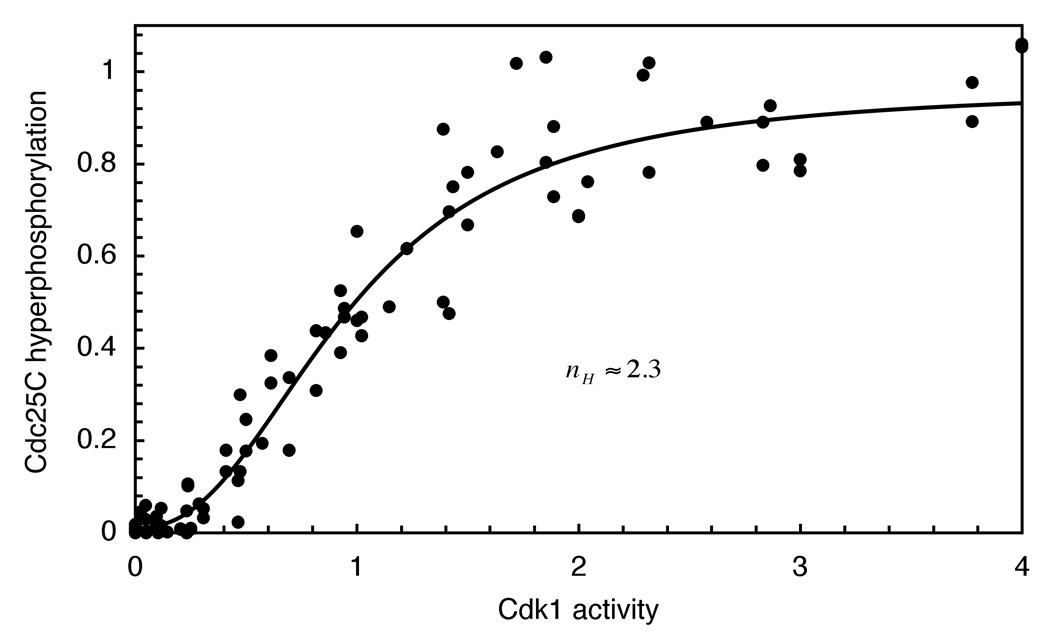
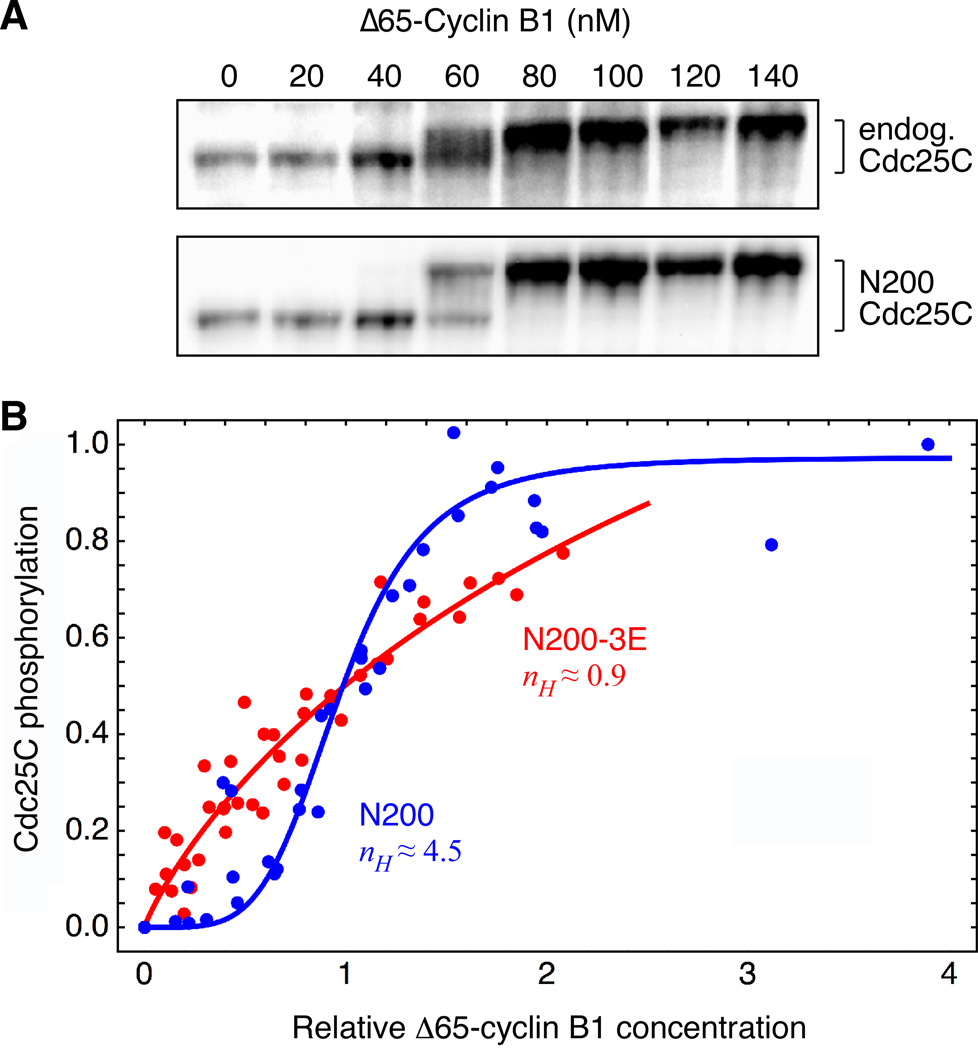
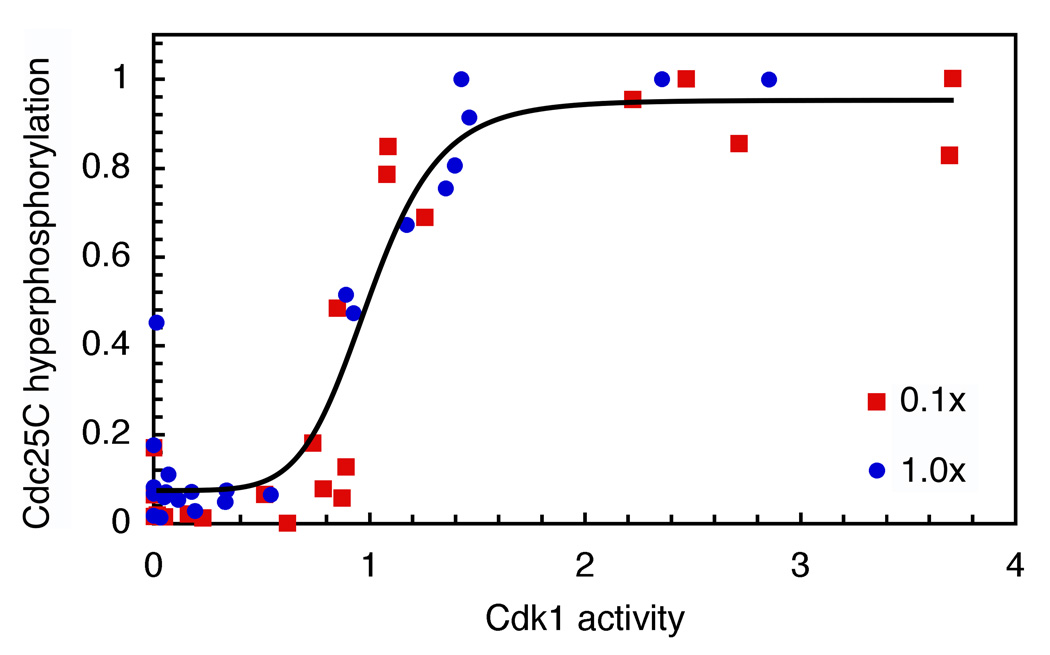
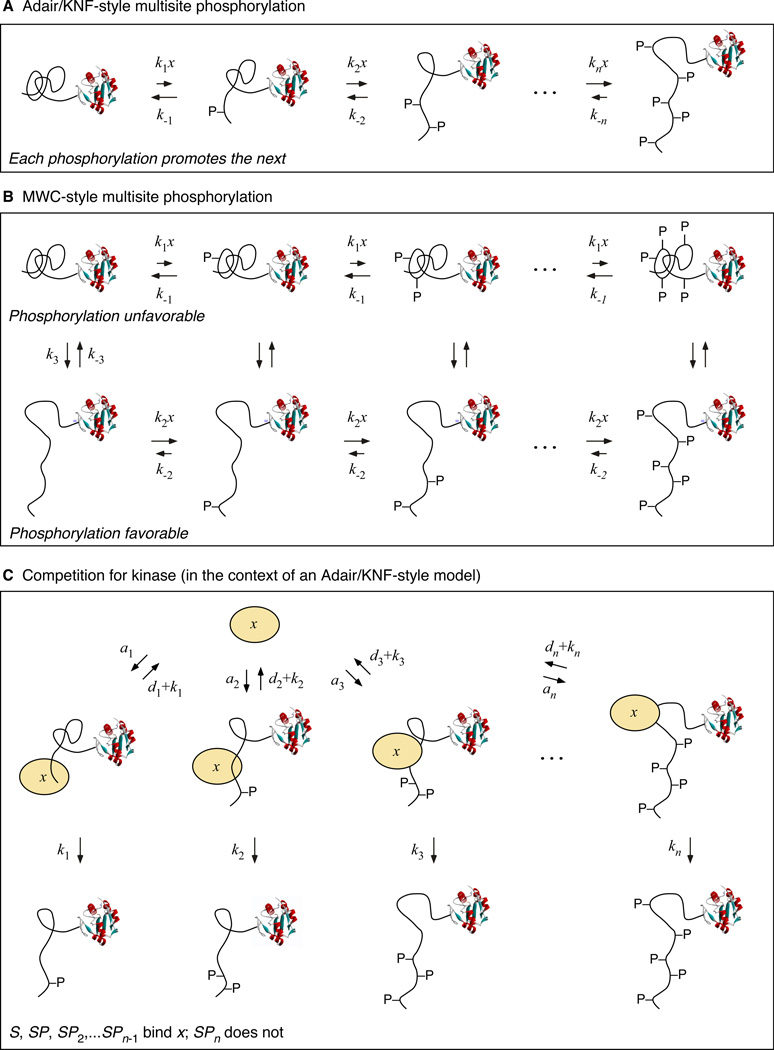
Cdc25C is a critical component of the interlinked positive and double-negative feedback loops that constitute the bistable mitotic trigger. Computational studies have indicated that the trigger's bistability should be more robust if the individual legs of the loops exhibit ultrasensitive responses. Here, we show that in Xenopus extracts two measures of Cdc25C activation (hyperphosphorylation and Ser 287 dephosphorylation) are highly ultrasensitive functions of the Cdk1 activity; estimated Hill coefficients were 11 to 32. Some of Cdc25C's ultrasensitivity can be reconstituted in vitro with purified components, and the reconstituted ultrasensitivity depends upon multisite phosphorylation. The response functions determined here for Cdc25C and previously for Wee1A allow us to formulate a simple mathematical model of the transition between interphase and mitosis. The model shows how the continuously variable regulators of mitosis work collectively to generate a switch-like, hysteretic response.
Copyright © 2011 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no financial conflicts of interest relevant to this work.
Figures
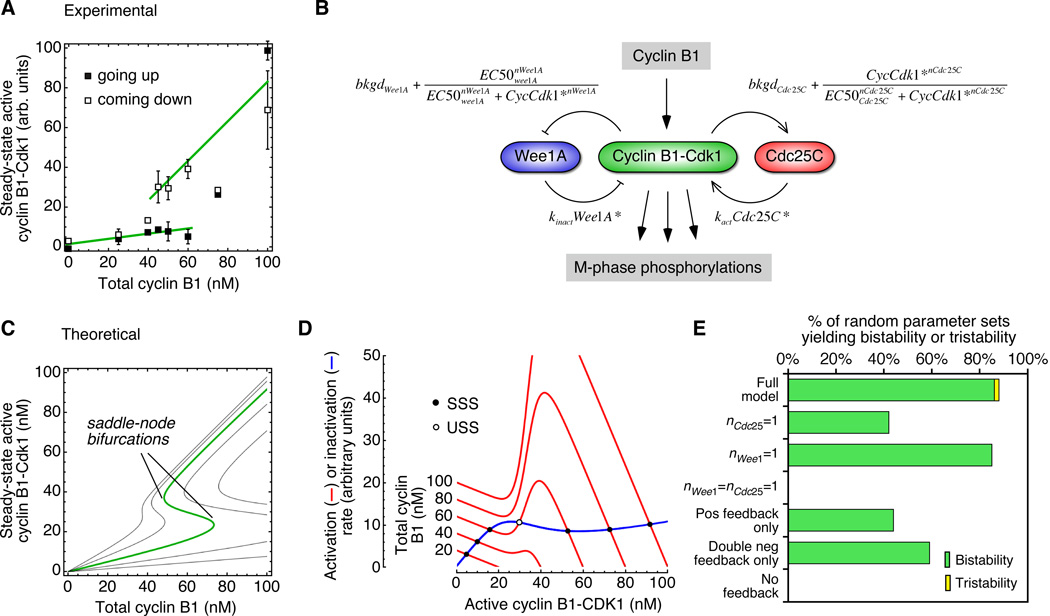
bkgdCdc25C = 0 to 0.4 (experimental estimate = 0.2)
bkgdWee1A = 0 to 0.4 (experimental estimate = 0.2)
EC50Cdc25C = 20 to 80 nM (experimental estimate = 30 nM)
EC50Wee1A = 20 to 80 nM (experimental estimate = 35 nM)
nCdc25C = 5 to 15 (experimental estimate = 11)
nWee1A = 1.5 to 8 (experimental estimate = 3.5)
kWee1A/kCdc25C = 0 to 1.5
Comment in
-
Ultrasensitivity and positive feedback to promote sharp mitotic entry.Mol Cell. 2011 Feb 4;41(3):243-4. doi: 10.1016/j.molcel.2011.01.016. Mol Cell. 2011. PMID: 21292155
Similar articles
-
Multisite phosphoregulation of Cdc25 activity refines the mitotic entrance and exit switches.Proc Natl Acad Sci U S A. 2012 Jun 19;109(25):9899-904. doi: 10.1073/pnas.1201366109. Epub 2012 Jun 4. Proc Natl Acad Sci U S A. 2012. PMID: 22665807 Free PMC article.
-
Control of mitotic exit by PP2A regulation of Cdc25C and Cdk1.Proc Natl Acad Sci U S A. 2007 Dec 11;104(50):19867-72. doi: 10.1073/pnas.0709879104. Epub 2007 Dec 4. Proc Natl Acad Sci U S A. 2007. PMID: 18056802 Free PMC article.
-
Bistable, Biphasic Regulation of PP2A-B55 Accounts for the Dynamics of Mitotic Substrate Phosphorylation.Curr Biol. 2021 Feb 22;31(4):794-808.e6. doi: 10.1016/j.cub.2020.11.058. Epub 2020 Dec 22. Curr Biol. 2021. PMID: 33357450 Free PMC article.
-
Switches and latches: a biochemical tug-of-war between the kinases and phosphatases that control mitosis.Philos Trans R Soc Lond B Biol Sci. 2011 Dec 27;366(1584):3584-94. doi: 10.1098/rstb.2011.0087. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 22084385 Free PMC article. Review.
-
Function and regulation of cdc25 protein phosphate through mitosis and meiosis.Prog Cell Cycle Res. 1995;1:215-28. doi: 10.1007/978-1-4615-1809-9_17. Prog Cell Cycle Res. 1995. PMID: 9552365 Review.
Cited by
-
Effect of magnitude and variability of energy of activation in multisite ultrasensitive biochemical processes.PLoS Comput Biol. 2020 Aug 6;16(8):e1007966. doi: 10.1371/journal.pcbi.1007966. eCollection 2020 Aug. PLoS Comput Biol. 2020. PMID: 32760072 Free PMC article.
-
Ultrasensitivity part II: multisite phosphorylation, stoichiometric inhibitors, and positive feedback.Trends Biochem Sci. 2014 Nov;39(11):556-69. doi: 10.1016/j.tibs.2014.09.003. Epub 2014 Oct 23. Trends Biochem Sci. 2014. PMID: 25440716 Free PMC article. Review.
-
MAP kinase modules: the excursion model and the steps that count.Biophys J. 2014 Nov 4;107(9):2006-15. doi: 10.1016/j.bpj.2014.09.024. Biophys J. 2014. PMID: 25418086 Free PMC article. Review.
-
Cyclin A regulates kinetochore microtubules to promote faithful chromosome segregation.Nature. 2013 Oct 3;502(7469):110-3. doi: 10.1038/nature12507. Epub 2013 Sep 8. Nature. 2013. PMID: 24013174 Free PMC article.
-
Ultrasensitivity part III: cascades, bistable switches, and oscillators.Trends Biochem Sci. 2014 Dec;39(12):612-8. doi: 10.1016/j.tibs.2014.10.002. Epub 2014 Nov 10. Trends Biochem Sci. 2014. PMID: 25456048 Free PMC article. Review.
References
-
- Adair GS. The hemoglobin system. VI. The oxygen dissociation curve of hemoglobin. J Biol Chem. 1925;63:529–545.
-
- Bonnet J, Coopman P, Morris MC. Characterization of centrosomal localization and dynamics of Cdc25C phosphatase in mitosis. Cell Cycle. 2008;7:1991–1998. - PubMed
-
- Booher RN, Holman PS, Fattaey A. Human Myt1 is a cell cycle-regulated kinase that inhibits Cdc2 but not Cdk2 activity. J Biol Chem. 1997;272:22300–22306. - PubMed
-
- Elia A, Cantley LC, Yaffe MB. Proteomic screen finds pSer/pThr-binding domain localizing Plk1 to mitotic substrates. Science. 2003;299:1228–1231. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
