

Phosphorylation dynamics regulate Hsp27-mediated rescue of neuronal plasticity deficits in tau transgenic mice
- PMID: 21084594
- PMCID: PMC3073547
- DOI: 10.1523/JNEUROSCI.3155-10.2010
Phosphorylation dynamics regulate Hsp27-mediated rescue of neuronal plasticity deficits in tau transgenic mice
Abstract
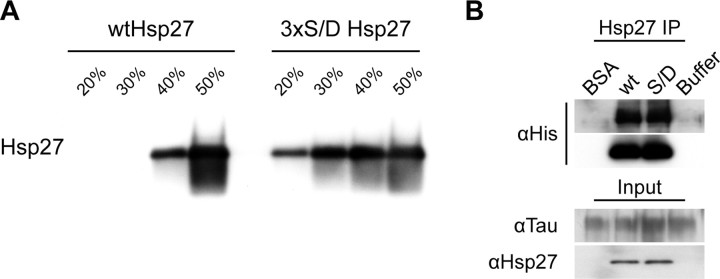
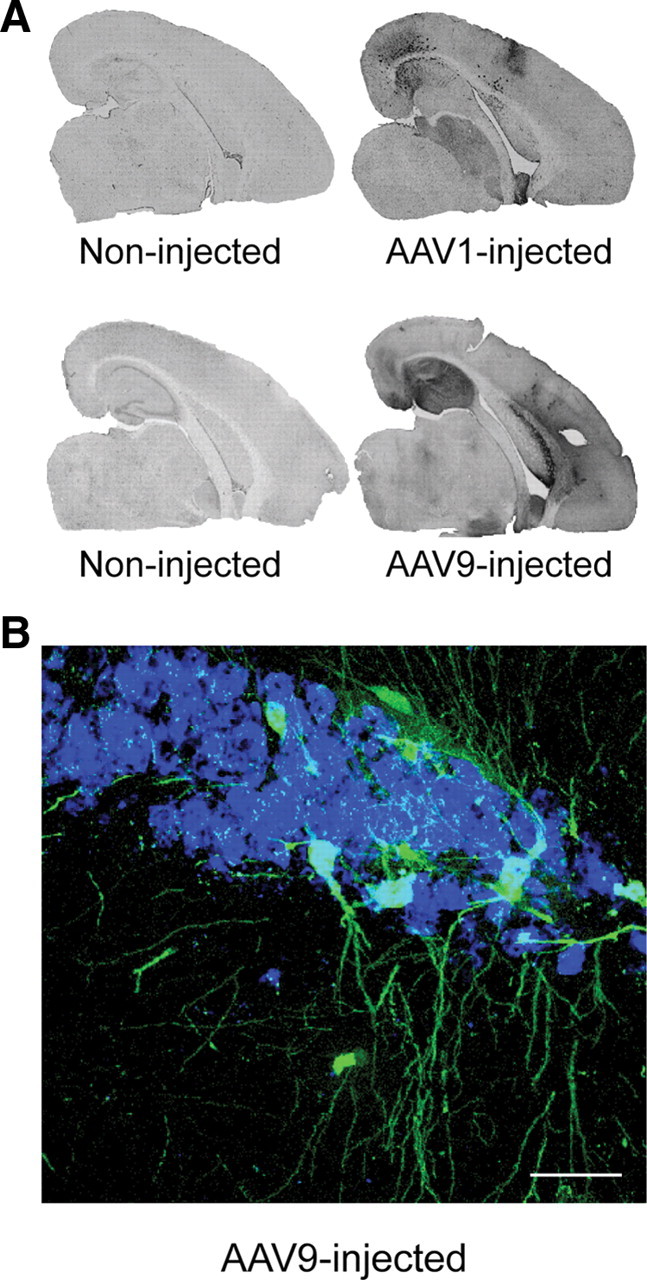
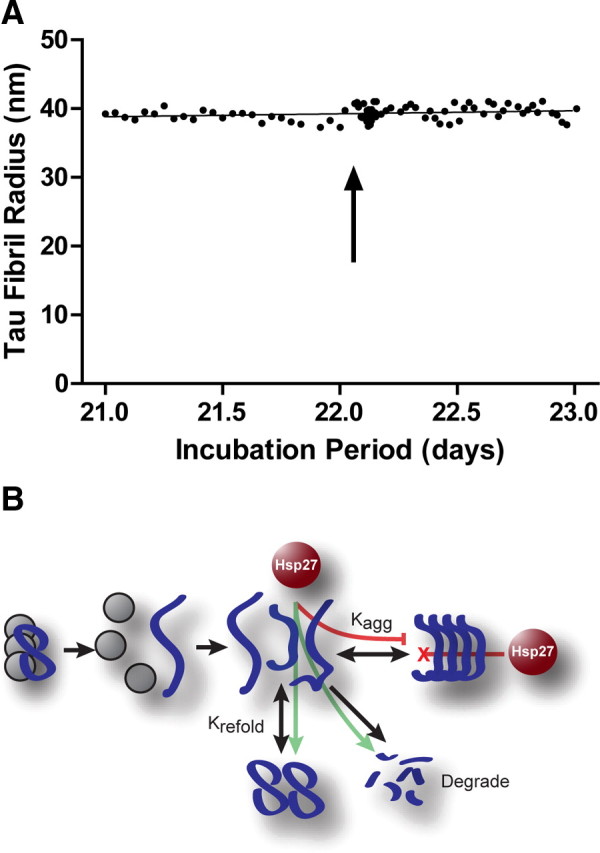
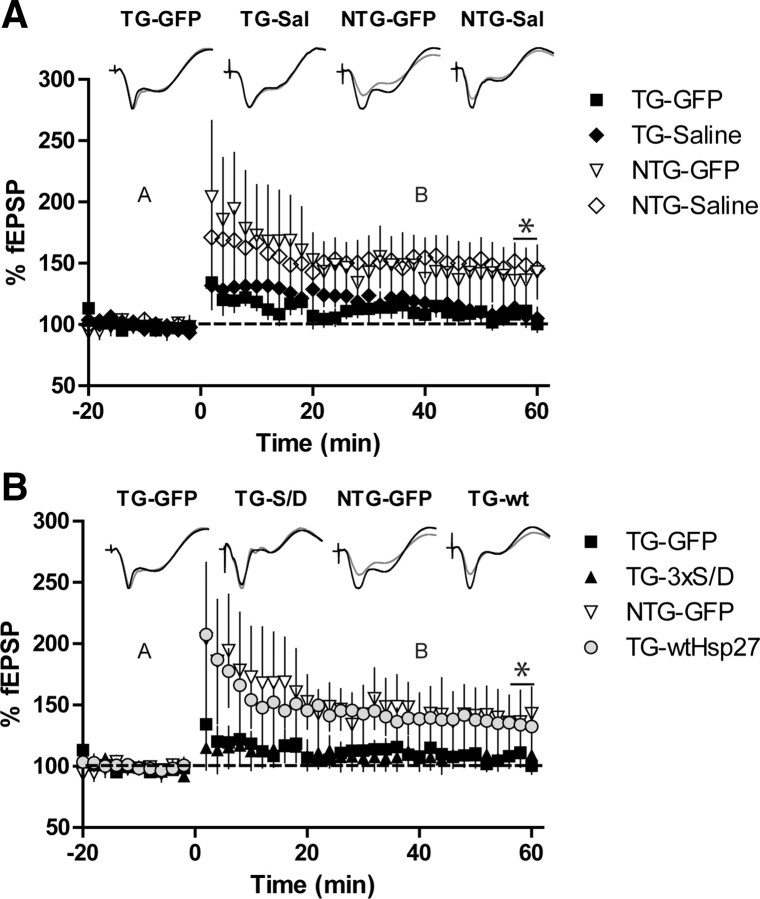
Molecular chaperones regulate the aggregation of a number of proteins that pathologically misfold and accumulate in neurodegenerative diseases. Identifying ways to manipulate these proteins in disease models is an area of intense investigation; however, the translation of these results to the mammalian brain has progressed more slowly. In this study, we investigated the ability of one of these chaperones, heat shock protein 27 (Hsp27), to modulate tau dynamics. Recombinant wild-type Hsp27 and a genetically altered version of Hsp27 that is perpetually pseudo-phosphorylated (3×S/D) were generated. Both Hsp27 variants interacted with tau, and atomic force microscopy and dynamic light scattering showed that both variants also prevented tau filament formation. However, extrinsic genetic delivery of these two Hsp27 variants to tau transgenic mice using adeno-associated viral particles showed that wild-type Hsp27 reduced neuronal tau levels, whereas 3×S/D Hsp27 was associated with increased tau levels. Moreover, rapid decay in hippocampal long-term potentiation (LTP) intrinsic to this tau transgenic model was rescued by wild-type Hsp27 overexpression but not by 3×S/D Hsp27. Because the 3×S/D Hsp27 mutant cannot cycle between phosphorylated and dephosphorylated states, we can conclude that Hsp27 must be functionally dynamic to facilitate tau clearance from the brain and rescue LTP; however, when this property is compromised, Hsp27 may actually facilitate accumulation of soluble tau intermediates.
Figures



Similar articles
-
Allosteric heat shock protein 70 inhibitors rapidly rescue synaptic plasticity deficits by reducing aberrant tau.Biol Psychiatry. 2013 Sep 1;74(5):367-74. doi: 10.1016/j.biopsych.2013.02.027. Epub 2013 Apr 19. Biol Psychiatry. 2013. PMID: 23607970 Free PMC article.
-
Small heat shock proteins Hsp27 or alphaB-crystallin and the protein components of neurofibrillary tangles: tau and neurofilaments.J Neurosci Res. 2008 May 1;86(6):1343-52. doi: 10.1002/jnr.21589. J Neurosci Res. 2008. PMID: 18061943
-
Interplay between glycogen synthase kinase-3β and tau in the cerebellum of Hsp27 transgenic mouse.J Neurosci Res. 2011 Aug;89(8):1267-75. doi: 10.1002/jnr.22660. Epub 2011 May 4. J Neurosci Res. 2011. PMID: 21544852
-
Tau phosphorylation, molecular chaperones, and ubiquitin E3 ligase: clinical relevance in Alzheimer's disease.J Alzheimers Dis. 2015;43(2):341-61. doi: 10.3233/JAD-140933. J Alzheimers Dis. 2015. PMID: 25096626 Review.
-
Phosphorylated tau and the neurodegenerative foldopathies.Biochim Biophys Acta. 2005 Jan 3;1739(2-3):298-310. doi: 10.1016/j.bbadis.2004.10.011. Epub 2004 Nov 26. Biochim Biophys Acta. 2005. PMID: 15615647 Review.
Cited by
-
Allosteric heat shock protein 70 inhibitors rapidly rescue synaptic plasticity deficits by reducing aberrant tau.Biol Psychiatry. 2013 Sep 1;74(5):367-74. doi: 10.1016/j.biopsych.2013.02.027. Epub 2013 Apr 19. Biol Psychiatry. 2013. PMID: 23607970 Free PMC article.
-
Effects of altered tau expression on dentate granule cell excitability in mice.Exp Neurol. 2021 Sep;343:113766. doi: 10.1016/j.expneurol.2021.113766. Epub 2021 May 21. Exp Neurol. 2021. PMID: 34029610 Free PMC article.
-
Mechanisms tailoring the expression of heat shock proteins to proteostasis challenges.J Biol Chem. 2022 May;298(5):101796. doi: 10.1016/j.jbc.2022.101796. Epub 2022 Mar 3. J Biol Chem. 2022. PMID: 35248532 Free PMC article. Review.
-
Tau protein aggregates inhibit the protein-folding and vesicular trafficking arms of the cellular proteostasis network.J Biol Chem. 2019 May 10;294(19):7917-7930. doi: 10.1074/jbc.RA119.007527. Epub 2019 Apr 1. J Biol Chem. 2019. PMID: 30936201 Free PMC article.
-
Small heat shock proteins in neurodegenerative diseases.Cell Stress Chaperones. 2020 Jul;25(4):679-699. doi: 10.1007/s12192-020-01101-4. Epub 2020 Apr 22. Cell Stress Chaperones. 2020. PMID: 32323160 Free PMC article. Review.
References
-
- Barghorn S, Davies P, Mandelkow E. Tau paired helical filaments from Alzheimer's disease brain and assembled in vitro are based on beta-structure in the core domain. Biochemistry. 2004;43:1694–1703. - PubMed
-
- Berrow NS, Büssow K, Coutard B, Diprose J, Ekberg M, Folkers GE, Levy N, Lieu V, Owens RJ, Peleg Y, Pinaglia C, Quevillon-Cheruel S, Salim L, Scheich C, Vincentelli R, Busso D. Recombinant protein expression and solubility screening in Escherichia coli: a comparative study. Acta Crystallogr D Biol Crystallogr. 2006;62:1218–1226. - PubMed
-
- Bolte S, Cordelières FP. A guided tour into subcellular colocalization analysis in light microscopy. J Microsc. 2006;224:213–232. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous