

doi: 10.1038/nm.2238.
Epub 2010 Oct 10.
Efficient hepatitis C virus particle formation requires diacylglycerol acyltransferase-1
Affiliations
- PMID: 20935628
- PMCID: PMC3431199
- DOI: 10.1038/nm.2238
Item in Clipboard
Efficient hepatitis C virus particle formation requires diacylglycerol acyltransferase-1
Nat Med.
2010 Nov.
Abstract
Hepatitis C virus (HCV) infection is closely tied to the lipid metabolism of liver cells. Here we identify the triglyceride-synthesizing enzyme diacylglycerol acyltransferase-1 (DGAT1) as a key host factor for HCV infection. DGAT1 interacts with the viral nucleocapsid core and is required for the trafficking of core to lipid droplets. Inhibition of DGAT1 activity or RNAi-mediated knockdown of DGAT1 severely impairs infectious virion production, implicating DGAT1 as a new target for antiviral therapy.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
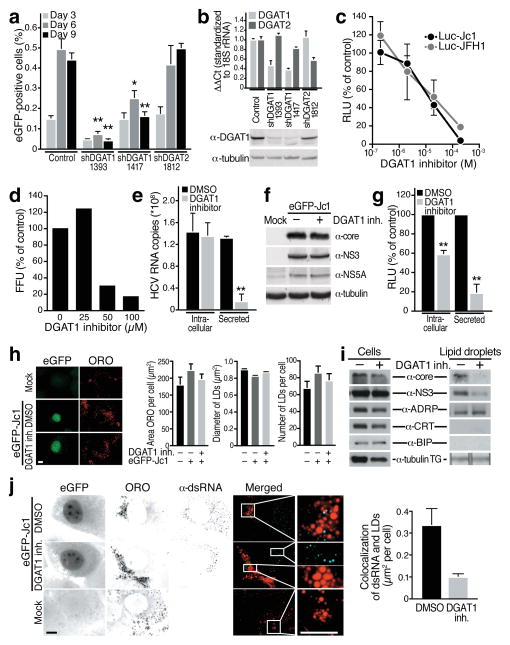
(a) Infection of shRNA-expressing Huh7.5 cells with low concentrations of eGFP-Jc1 viral stock. Spreading viral infection was measured by flow cytometry of eGFP (mean ± s.e.m.; n = 4; *p < 0.05, **p < 0.01). (b) Real-time RT-PCR analysis of DGAT1 or DGAT2 mRNAs in knockdown cells (mean ± s.e.m.; n = 4) or western blot analysis of DGAT1 protein expression. Available antibodies against DGAT2 do not reliably detect endogenous human DGAT2 in our hands. (c) Dose-dependent decrease of infectious titres in Huh7.5 cells transfected with Luciferase-Jc1 or Luciferase-JFH1 RNA and incubated with increasing concentrations of the DGAT1 inhibitor or DMSO for 48 h. Naïve Huh7.5 cells were infected with cell supernatants of treated cells and lysed 48 h post-infection to analyze luciferase activity (expressed as percent relative to DMSO control; mean ± s.e.m.: n = 3). (d) Dose-dependent decrease of infectious virus titers released from freshly isolated primary human hepatocytes infected with HCV-Jc1 viral stock and treated with increasing amounts of the DGAT1 inhibitor or DMSO for 3 days. Shown are infectivity titers (in FFU) expressed as percent of DMSO control. A single experiment is shown. (e) Real-time RT-PCR analysis of HCV RNA isolated from cells or from supernatants of Huh7.5 cells electroporated with eGFP-Jc1 RNA and treated with DGAT1 inhibitor (20 μM) or DMSO. Results are expressed as HCV RNA copy numbers per 1 μg total cellular RNA normalized to 18S rRNA (Intracellular) or per 1 ml culture supernatant (Secreted) at day 4 after transfection (mean ± s.d.; n = 6; **p < 0.01). (f) Western blot analysis of cell extracts described in e) lysed at day 4 after transfection. (g) Infection of naïve Huh7.5 cells with either intracellular or secreted viral particles isolated from Huh7.5 transfected with Luciferase-Jc1 RNA and treated with the DGAT1 inhibitor (20 μM) or DMSO. Shown are luciferase values expressed as percent of DMSO control (mean ± s.d.; n = 3; **p < 0.01). (h) Representative images and quantification of epifluorescence of Huh7 Lunet cells electroporated with eGFP-Jc1 RNA and treated with DMSO or DGAT1 inhibitor (20 μM) after oil-red-O (ORO) staining. LD area: mean of 1000 cells ± s.e.m.; LD diameter: mean of > 1600 LDs ± s.e.m.; LD number: mean of > 50 cells ± s.e.m. (i) Western blot analysis of cell extracts or isolated LD fractions from cells described in h). TG: extracted triglycerides analyzed by thin layer chromatography. (j) Indirect immunofluorescence of double-stranded RNA (α-dsRNA) at LDs (ORO) in cells described in h) (scale bar = 10 μm) and quantification of overlap of the signals for double-stranded RNAs and ORO per cell (mean of 30 cells ± s.e.m.).
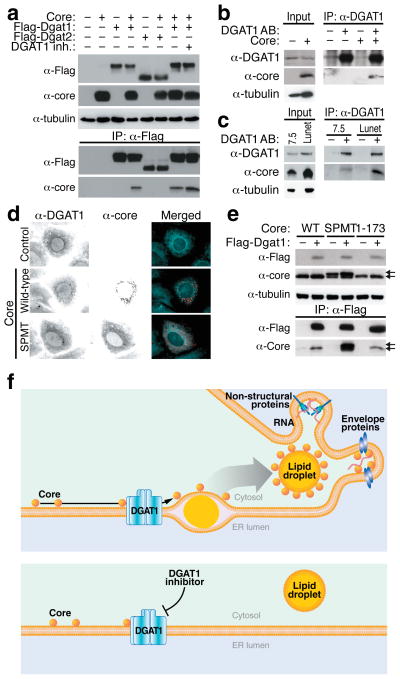
(a) Coimmunoprecipitation assays in 293T cells transfected with expression vectors for the HCV core protein and Flag-DGAT1 or Flag-DGAT2 proteins and treated with DGAT1 inhibitor (20 μM) or DMSO. After immunoprecipitation with α-Flag agarose, the core protein was detected by western blotting with α-core antibodies. (b) Coimmunoprecipitation of the HCV core protein with endogenous DGAT1 in Huh7 hepatoma cells transduced with a core-expressing lentiviral vector. (c) Coimmunoprecipitation of the HCV core protein with endogenous DGAT1 in Huh7.5 and Huh7 Lunet cells electroporated with eGFP-Jc1 RNA. (d) Indirect immunofluorescence of core and endogenous DGAT1 in Huh7 cells transfected with wild-type or mutant (SPMT) core expression vectors (scale bar = 10 μm). (e) Coimmunoprecipitation assays in 293T cells transfected with expression vectors for wild-type (WT), mutant (SPMT) or truncated (1–173) core protein and the Flag-DGAT1-expressing plasmid. Arrows mark unprocessed (upper) and processed (lower) core protein. (f) Model of HCV core recruitment to DGAT1-generated LDs. See text for details.
Comment in
-
Diacylglycerol acyltransferase-1: a critical host factor for hepatitis C virus assembly and potential new drug target.Gastroenterology. 2011 Apr;140(4):1345-7. doi: 10.1053/j.gastro.2011.02.037. Epub 2011 Feb 24. Gastroenterology. 2011. PMID: 21354164 No abstract available.
-
Triglyceride synthesis and hepatitis C virus production: identification of a novel host factor as antiviral target.Hepatology. 2011 Mar;53(3):1046-8. doi: 10.1002/hep.24177. Hepatology. 2011. PMID: 21374668 No abstract available.
Similar articles
-
The triglyceride-synthesizing enzyme diacylglycerol acyltransferase 2 modulates the formation of the hepatitis C virus replication organelle.PLoS Pathog. 2024 Sep 6;20(9):e1012509. doi: 10.1371/journal.ppat.1012509. eCollection 2024 Sep. PLoS Pathog. 2024. PMID: 39241103 Free PMC article.
-
Cortactin Interacts with Hepatitis C Virus Core and NS5A Proteins: Implications for Virion Assembly.J Virol. 2020 Sep 15;94(19):e01306-20. doi: 10.1128/JVI.01306-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32727880 Free PMC article.
-
Hepatitis C virus entry is impaired by claudin-1 downregulation in diacylglycerol acyltransferase-1-deficient cells.J Virol. 2014 Aug;88(16):9233-44. doi: 10.1128/JVI.01428-14. Epub 2014 Jun 4. J Virol. 2014. PMID: 24899196 Free PMC article.
-
Assembly of hepatitis C virus particles.Microbiol Immunol. 2011 Jan;55(1):12-8. doi: 10.1111/j.1348-0421.2010.00274.x. Microbiol Immunol. 2011. PMID: 21175769 Review.
-
Current status of the research and development of diacylglycerol O-acyltransferase 1 (DGAT1) inhibitors.J Med Chem. 2013 Dec 27;56(24):9820-5. doi: 10.1021/jm4007033. Epub 2013 Sep 3. J Med Chem. 2013. PMID: 23919406 Review.
Cited by
-
Infections at the nexus of metabolic-associated fatty liver disease.Arch Toxicol. 2021 Jul;95(7):2235-2253. doi: 10.1007/s00204-021-03069-1. Epub 2021 May 24. Arch Toxicol. 2021. PMID: 34027561 Free PMC article. Review.
-
Glycerophospholipid remodeling is critical for orthoflavivirus infection.Nat Commun. 2024 Oct 7;15(1):8683. doi: 10.1038/s41467-024-52979-y. Nat Commun. 2024. PMID: 39375358 Free PMC article.
-
The Dynamic Landscape of Capsid Proteins and Viral RNA Interactions in Flavivirus Genome Packaging and Virus Assembly.Pathogens. 2024 Jan 28;13(2):120. doi: 10.3390/pathogens13020120. Pathogens. 2024. PMID: 38392858 Free PMC article. Review.
-
Virion assembly and release.Curr Top Microbiol Immunol. 2013;369:199-218. doi: 10.1007/978-3-642-27340-7_8. Curr Top Microbiol Immunol. 2013. PMID: 23463202 Free PMC article. Review.
-
Landscape of protein-protein interactions during hepatitis C virus assembly and release.Microbiol Spectr. 2024 Feb 6;12(2):e0256222. doi: 10.1128/spectrum.02562-22. Epub 2024 Jan 17. Microbiol Spectr. 2024. PMID: 38230952 Free PMC article.
References
-
- Moradpour D, Penin F, Rice CM. Nat Rev Microbiol. 2007;5:453–463. - PubMed
-
- Miyanari Y, et al. Nat Cell Biol. 2007;9:1089–1097. - PubMed
-
- Roingeard P, Hourioux C, Blanchard E, Prensier G. Histochem Cell Biol. 2008;130:561–566. - PubMed
-
- Boulant S, Targett-Adams P, McLauchlan J. J Gen Virol. 2007;88:2204–2213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
