

PGC-1α, a potential therapeutic target for early intervention in Parkinson's disease
- PMID: 20926834
- PMCID: PMC3129986
- DOI: 10.1126/scitranslmed.3001059
PGC-1α, a potential therapeutic target for early intervention in Parkinson's disease
Abstract
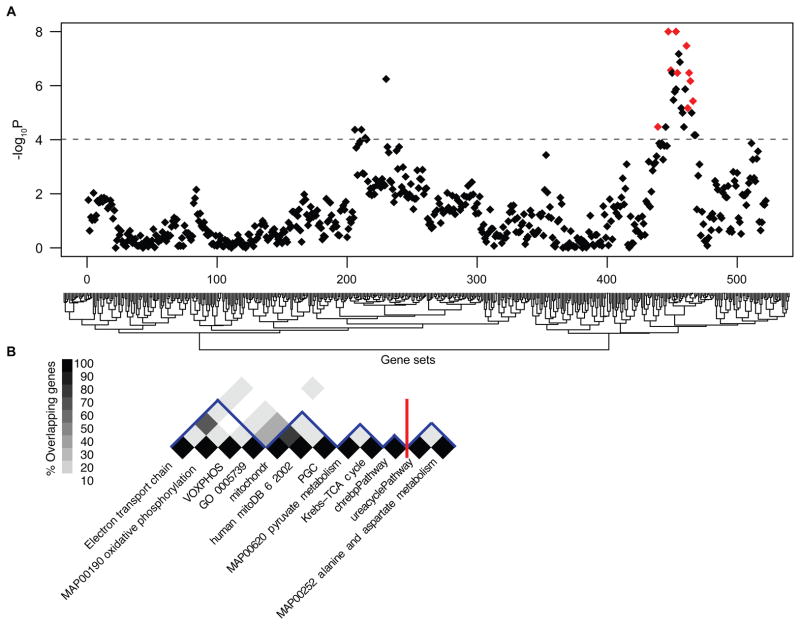
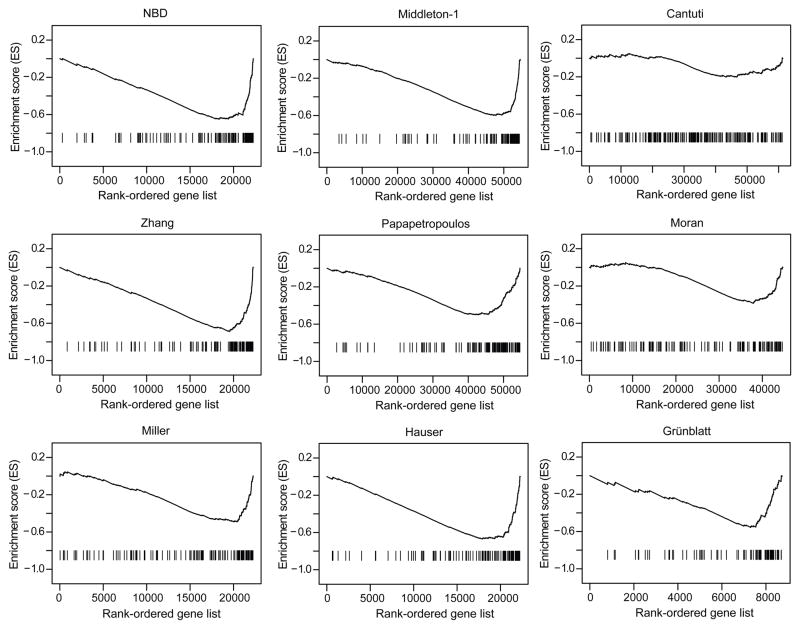
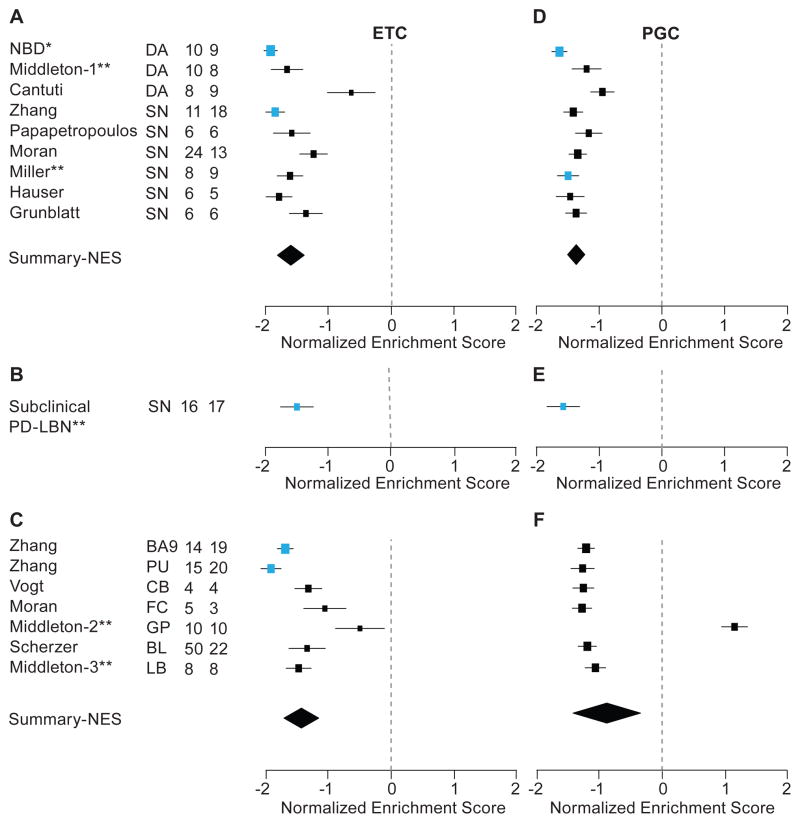
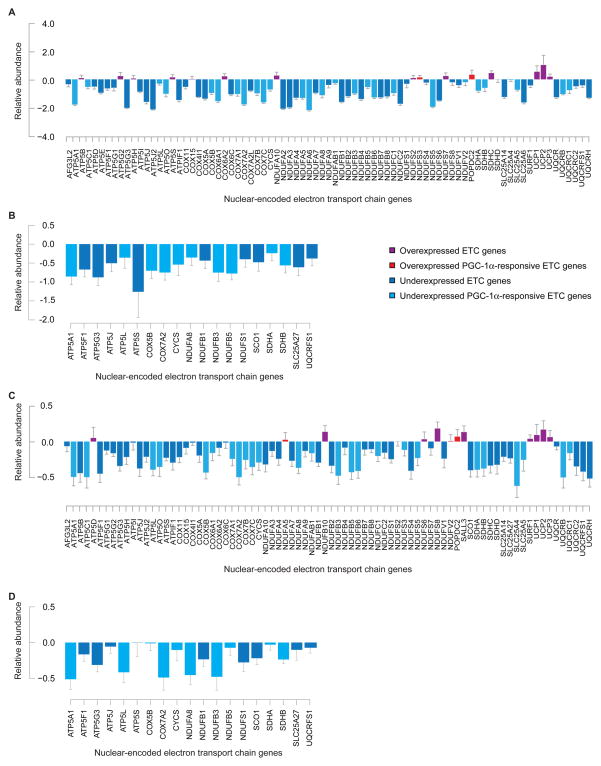
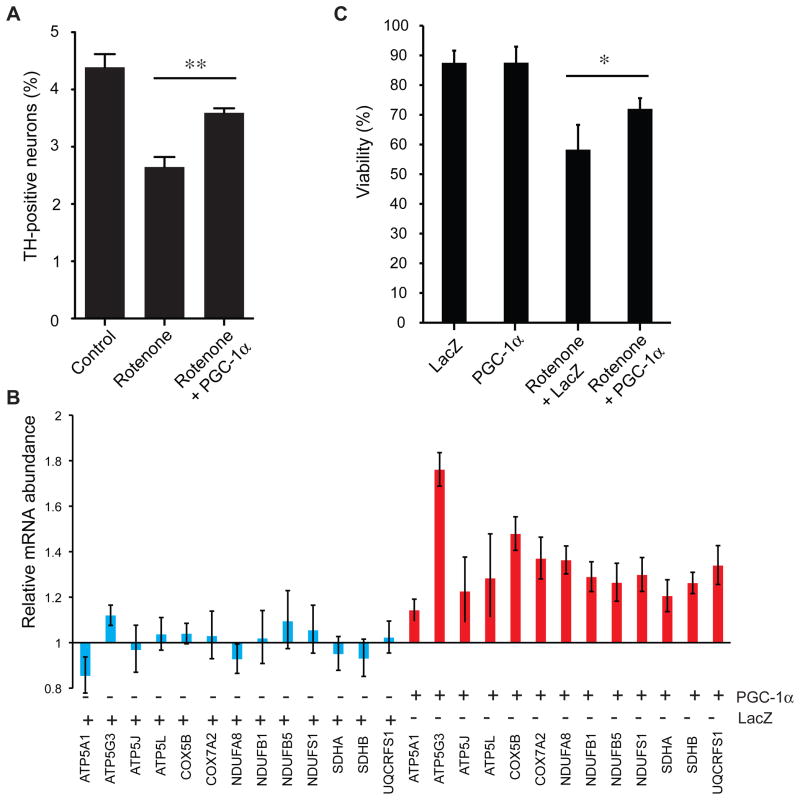
Parkinson's disease affects 5 million people worldwide, but the molecular mechanisms underlying its pathogenesis are still unclear. Here, we report a genome-wide meta-analysis of gene sets (groups of genes that encode the same biological pathway or process) in 410 samples from patients with symptomatic Parkinson's and subclinical disease and healthy controls. We analyzed 6.8 million raw data points from nine genome-wide expression studies, and 185 laser-captured human dopaminergic neuron and substantia nigra transcriptomes, followed by two-stage replication on three platforms. We found 10 gene sets with previously unknown associations with Parkinson's disease. These gene sets pinpoint defects in mitochondrial electron transport, glucose utilization, and glucose sensing and reveal that they occur early in disease pathogenesis. Genes controlling cellular bioenergetics that are expressed in response to peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) are underexpressed in Parkinson's disease patients. Activation of PGC-1α results in increased expression of nuclear-encoded subunits of the mitochondrial respiratory chain and blocks the dopaminergic neuron loss induced by mutant α-synuclein or the pesticide rotenone in cellular disease models. Our systems biology analysis of Parkinson's disease identifies PGC-1α as a potential therapeutic target for early intervention.
Conflict of interest statement
Figures

Similar articles
-
Brain-derived and in vitro-seeded alpha-synuclein fibrils exhibit distinct biophysical profiles.Elife. 2024 Nov 25;13:RP92775. doi: 10.7554/eLife.92775. Elife. 2024. PMID: 39584804 Free PMC article.
-
Medications for chronic obstructive pulmonary disease: a historical non-interventional cohort study with validation against RCT results.Health Technol Assess. 2021 Aug;25(51):1-70. doi: 10.3310/hta25510. Health Technol Assess. 2021. PMID: 34463610 Clinical Trial.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Dopamine and reward hypersensitivity in Parkinson's disease with impulse control disorder.Brain. 2020 Aug 1;143(8):2502-2518. doi: 10.1093/brain/awaa198. Brain. 2020. PMID: 32761061 Free PMC article.
-
The effectiveness of abstinence-based and harm reduction-based interventions in reducing problematic substance use in adults who are experiencing homelessness in high income countries: A systematic review and meta-analysis: A systematic review.Campbell Syst Rev. 2024 Apr 21;20(2):e1396. doi: 10.1002/cl2.1396. eCollection 2024 Jun. Campbell Syst Rev. 2024. PMID: 38645303 Free PMC article. Review.
Cited by
-
Targeting the Mitochondria-Proteostasis Axis to Delay Aging.Front Cell Dev Biol. 2021 Mar 11;9:656201. doi: 10.3389/fcell.2021.656201. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33777963 Free PMC article.
-
Exogenous Alpha-Synuclein Evoked Parkin Downregulation Promotes Mitochondrial Dysfunction in Neuronal Cells. Implications for Parkinson's Disease Pathology.Front Aging Neurosci. 2021 Feb 24;13:591475. doi: 10.3389/fnagi.2021.591475. eCollection 2021. Front Aging Neurosci. 2021. PMID: 33716707 Free PMC article.
-
Parkin in the regulation of fat uptake and mitochondrial biology: emerging links in the pathophysiology of Parkinson's disease.Curr Opin Lipidol. 2012 Jun;23(3):201-205. doi: 10.1097/MOL.0b013e328352dc5d. Curr Opin Lipidol. 2012. PMID: 22488424 Free PMC article. Review.
-
Delta-9-tetrahydrocannabinol protects against MPP+ toxicity in SH-SY5Y cells by restoring proteins involved in mitochondrial biogenesis.Oncotarget. 2016 Jul 19;7(29):46603-46614. doi: 10.18632/oncotarget.10314. Oncotarget. 2016. PMID: 27366949 Free PMC article.
-
Transcriptional pathobiology and multi-omics predictors for Parkinson's disease.bioRxiv [Preprint]. 2024 Jun 21:2024.06.18.599639. doi: 10.1101/2024.06.18.599639. bioRxiv. 2024. PMID: 38948706 Free PMC article. Preprint.
References
-
- Dhillon AS, Tarbutton GL, Levin JL, Plotkin GM, Lowry LK, Nalbone JT, Shepherd S. Pesticide/environmental exposures and Parkinson’s disease in East Texas. J Agromedicine. 2008;13:37. - PubMed
-
- Chade AR, Kasten M, Tanner CM. Nongenetic causes of Parkinson’s disease. J Neural Transm. 2006;(Suppl):147. - PubMed
-
- Park HJ, Kim HJ, Park HK, Chung JH. Protective effect of histamine H2 receptor antagonist ranitidine against rotenone-induced apoptosis. Neurotoxicology. 2009 Nov;30:1114. - PubMed
-
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat Neurosci. 2000 Dec;3:1301. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- P01 NS058793/NS/NINDS NIH HHS/United States
- R01 NS064155/NS/NINDS NIH HHS/United States
- P30AG19610/AG/NIA NIH HHS/United States
- R24MH068855/MH/NIMH NIH HHS/United States
- R01NS049221/NS/NINDS NIH HHS/United States
- K08 AG024816/AG/NIA NIH HHS/United States
- R21NS060227/NS/NINDS NIH HHS/United States
- P01NS058793/NS/NINDS NIH HHS/United States
- R01 NS049221-02/NS/NINDS NIH HHS/United States
- K08AG024816/AG/NIA NIH HHS/United States
- P30 AG019610/AG/NIA NIH HHS/United States
- R01 NS049221/NS/NINDS NIH HHS/United States
- P30 AG019610-01/AG/NIA NIH HHS/United States
- R01NS064155/NS/NINDS NIH HHS/United States
- R24 MH068855-01/MH/NIMH NIH HHS/United States
- R21 NS060227-01/NS/NINDS NIH HHS/United States
- P01 NS058793-02/NS/NINDS NIH HHS/United States
- R01 NS064155-01/NS/NINDS NIH HHS/United States
- R21 NS060227/NS/NINDS NIH HHS/United States
- R01 NS064155-02S1/NS/NINDS NIH HHS/United States
- K08 AG024816-05/AG/NIA NIH HHS/United States
- R24 MH068855/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
