

Endocytosis and intracellular trafficking of Notch and its ligands
- PMID: 20816395
- PMCID: PMC6233319
- DOI: 10.1016/S0070-2153(10)92005-X
Endocytosis and intracellular trafficking of Notch and its ligands
Abstract
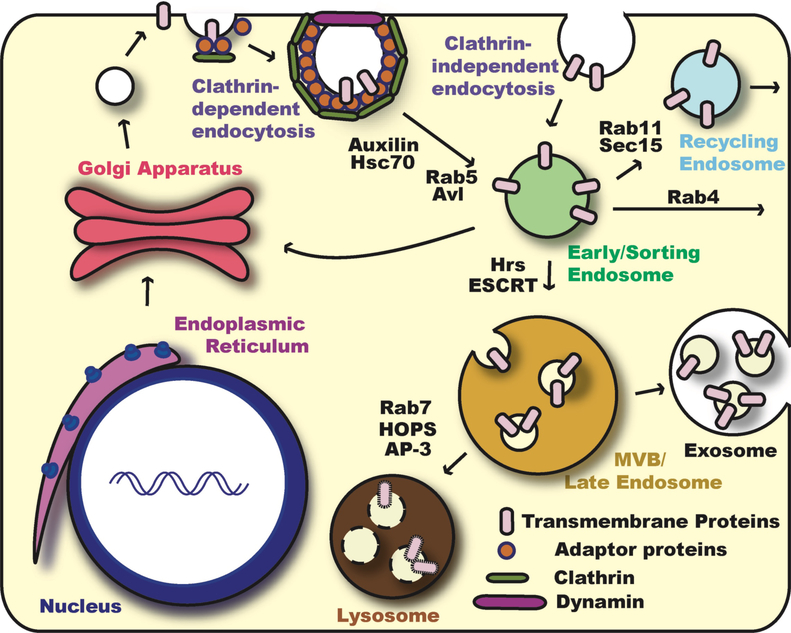
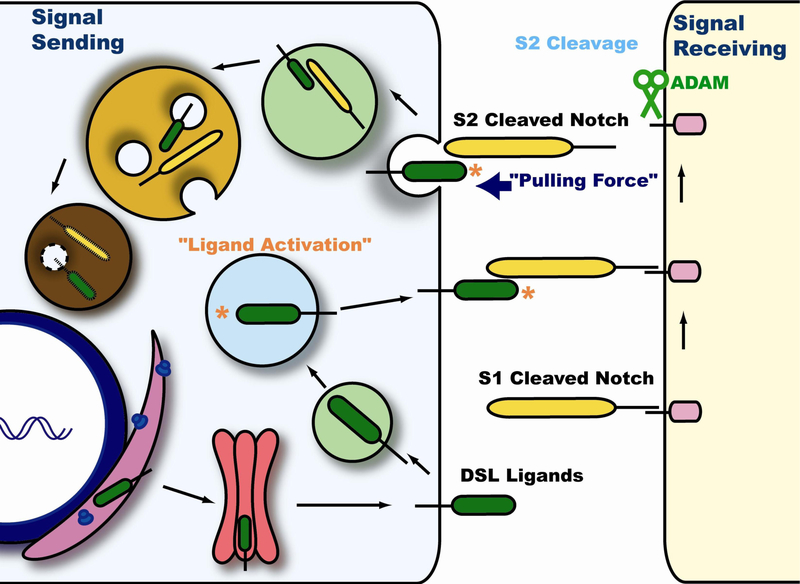
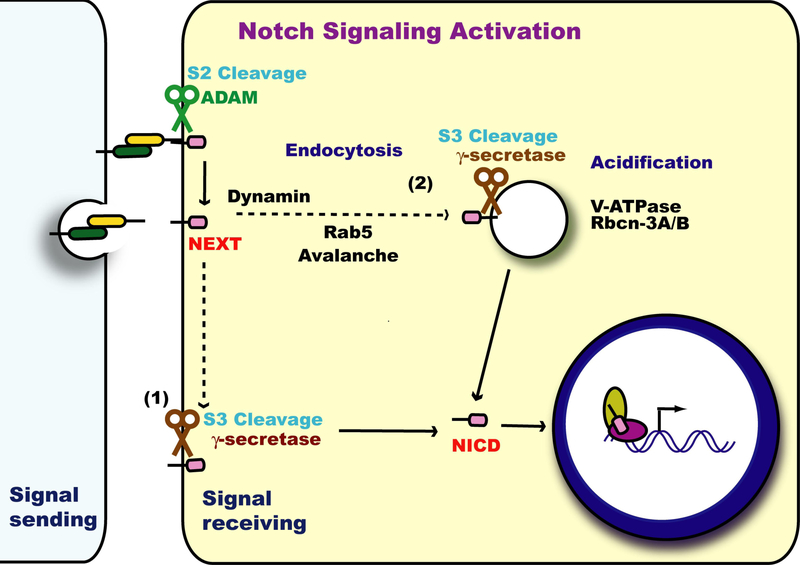
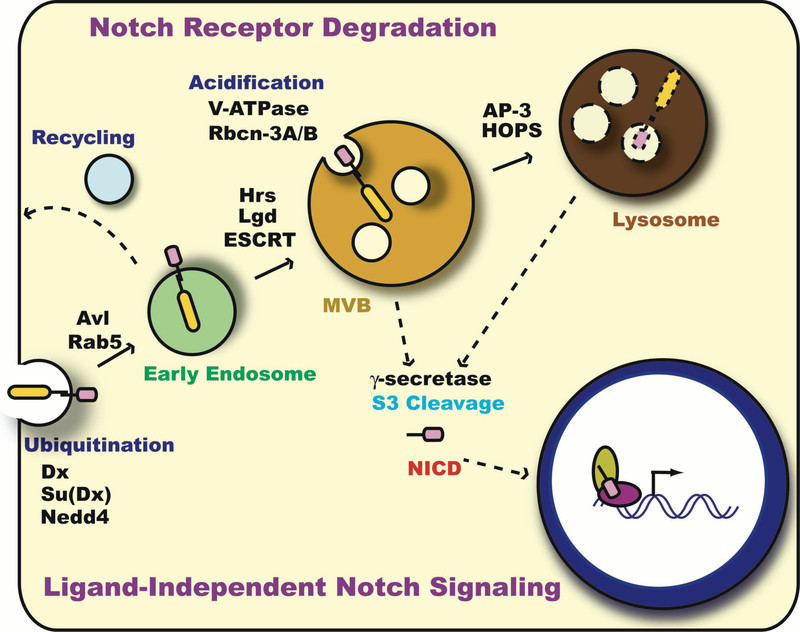
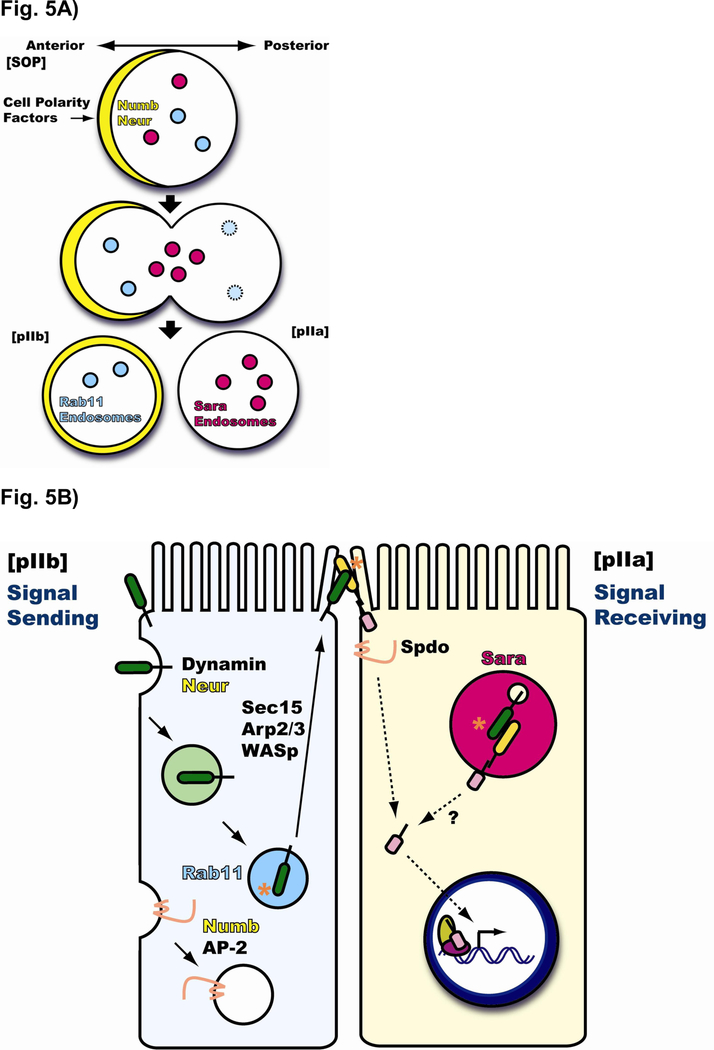
Notch signaling occurs through direct interaction between Notch, the receptor, and its ligands, presented on the surface of neighboring cells. Endocytosis has been shown to be essential for Notch signal activation in both signal-sending and signal-receiving cells, and numerous genes involved in vesicle trafficking have recently been shown to act as key regulators of the pathway. Defects in vesicle trafficking can lead to gain- or loss-of-function defects in a context-dependent manner. Here, we discuss how endocytosis and vesicle trafficking regulate Notch signaling in both signal-sending and signal-receiving cells. We will introduce the key players in different trafficking steps, and further illustrate how they impact the signal outcome. Some of these players act as general factors and modulate Notch signaling in all contexts, whereas others modulate signaling in a context-specific fashion. We also discuss Notch signaling during mechanosensory organ development in the fly to exemplify how endocytosis and vesicle trafficking are effectively used to determine correct cell fates. In summary, endocytosis plays an essential role in Notch signaling, whereas intracellular vesicle trafficking often plays a context-dependent or regulatory role, leading to divergent outcomes in different developmental contexts.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
The cell giveth and the cell taketh away: an overview of Notch pathway activation by endocytic trafficking of ligands and receptors.Acta Histochem. 2011 May;113(3):248-55. doi: 10.1016/j.acthis.2010.01.006. Epub 2010 Feb 1. Acta Histochem. 2011. PMID: 20122714 Free PMC article. Review.
-
Notch receptor-ligand interactions during T cell development, a ligand endocytosis-driven mechanism.Curr Top Microbiol Immunol. 2012;360:19-46. doi: 10.1007/82_2012_225. Curr Top Microbiol Immunol. 2012. PMID: 22581027 Review.
-
Endocytic regulation of Notch signaling.Curr Opin Genet Dev. 2009 Aug;19(4):323-8. doi: 10.1016/j.gde.2009.04.005. Epub 2009 May 15. Curr Opin Genet Dev. 2009. PMID: 19447603 Free PMC article. Review.
-
Genetic identification of intracellular trafficking regulators involved in Notch-dependent binary cell fate acquisition following asymmetric cell division.J Cell Sci. 2012 Oct 15;125(Pt 20):4886-901. doi: 10.1242/jcs.110171. Epub 2012 Jul 23. J Cell Sci. 2012. PMID: 22825875
-
Endocytosis and control of Notch signaling.Curr Opin Cell Biol. 2012 Aug;24(4):534-40. doi: 10.1016/j.ceb.2012.06.006. Epub 2012 Jul 18. Curr Opin Cell Biol. 2012. PMID: 22818956 Free PMC article. Review.
Cited by
-
The lack of autophagy triggers precocious activation of Notch signaling during Drosophila oogenesis.BMC Dev Biol. 2012 Dec 5;12:35. doi: 10.1186/1471-213X-12-35. BMC Dev Biol. 2012. PMID: 23217079 Free PMC article.
-
BLOS2 negatively regulates Notch signaling during neural and hematopoietic stem and progenitor cell development.Elife. 2016 Oct 10;5:e18108. doi: 10.7554/eLife.18108. Elife. 2016. PMID: 27719760 Free PMC article.
-
Disruption of Drosophila melanogaster lipid metabolism genes causes tissue overgrowth associated with altered developmental signaling.PLoS Genet. 2013 Nov;9(11):e1003917. doi: 10.1371/journal.pgen.1003917. Epub 2013 Nov 7. PLoS Genet. 2013. PMID: 24244188 Free PMC article.
-
Notch3 pathway alterations in ovarian cancer.Cancer Res. 2014 Jun 15;74(12):3282-93. doi: 10.1158/0008-5472.CAN-13-2066. Epub 2014 Apr 17. Cancer Res. 2014. PMID: 24743243 Free PMC article.
-
The adaptor-associated kinase 1, AAK1, is a positive regulator of the Notch pathway.J Biol Chem. 2011 May 27;286(21):18720-30. doi: 10.1074/jbc.M110.190769. Epub 2011 Apr 4. J Biol Chem. 2011. PMID: 21464124 Free PMC article.
References
-
- Acconcia F, Sigismund S, and Polo S (2009). Ubiquitin in trafficking: the network at work. Experimental cell research 315, 1610–1618. - PubMed
-
- Bardin AJ, Le Borgne R, and Schweisguth F (2004). Asymmetric localization and function of cell-fate determinants: a fly’s view. Current opinion in neurobiology 14, 6–14. - PubMed
-
- Bardin AJ, and Schweisguth F (2006). Bearded family members inhibit Neuralized-mediated endocytosis and signaling activity of Delta in Drosophila. Developmental cell 10, 245–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
