

Identification of dendritic antigen-presenting cells in the zebrafish
- PMID: 20733076
- PMCID: PMC2936643
- DOI: 10.1073/pnas.1000494107
Identification of dendritic antigen-presenting cells in the zebrafish
Abstract
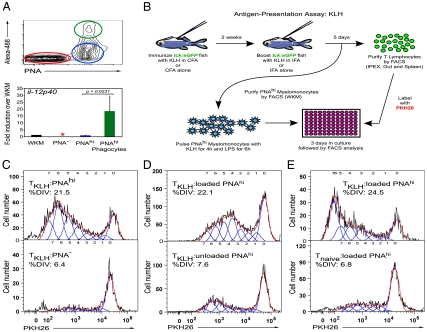
In mammals, dendritic cells (DCs) form the key link between the innate and adaptive immune systems. DCs act as immune sentries in various tissues and, upon encountering pathogen, engulf and traffic foreign antigen to secondary lymphoid tissues, stimulating antigen-specific T lymphocytes. Although DCs are of fundamental importance in orchestrating the mammalian immune response, their presence and function in nonmammalian vertebrates is largely unknown. Because teleosts possess one of the earliest recognizable adaptive immune systems, we sought to identify antigen-presenting cells (APCs) in the zebrafish to better understand the potential origins of DCs and their evolutionary relationship to lymphocytes. Here we present the identification and characterization of a zebrafish APC subset strongly resembling mammalian DCs. Rare DCs are present in various adult tissues, and can be enriched by their affinity for the lectin peanut agglutinin (PNA). We show that PNA(hi) myeloid cells possess the classical morphological features of mammalian DCs as revealed by histochemical and ultrastructural analyses, phagocytose-labeled bacterial preparations in vivo, and exhibit expression of genes associated with DC function and antigen presentation, including il12, MHC class II invariant chain iclp1, and csf1r. Importantly, we show that PNA(hi) cells can activate T lymphocytes in an antigen-dependent manner. Together, these studies suggest that the cellular constituents responsible for antigen presentation are remarkably conserved from teleosts to mammals, and indicate that the zebrafish may serve as a unique model to study the origin of APC subsets and their evolutionary role as the link between the innate and adaptive immune systems.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Dendritic type, accessory cells within the mammalian thymic microenvironment. Antigen presentation in the dendritic neuro-endocrine-immune cellular network.In Vivo. 1997 Jul-Aug;11(4):351-70. In Vivo. 1997. PMID: 9292303
-
Characterization of surface phenotypic molecules of teleost dendritic cells.Dev Comp Immunol. 2015 Mar;49(1):38-43. doi: 10.1016/j.dci.2014.11.010. Epub 2014 Nov 13. Dev Comp Immunol. 2015. PMID: 25445909
-
Perspectives on antigen presenting cells in zebrafish.Dev Comp Immunol. 2014 Sep;46(1):63-73. doi: 10.1016/j.dci.2014.03.010. Epub 2014 Mar 29. Dev Comp Immunol. 2014. PMID: 24685511 Free PMC article. Review.
-
Identification of Barramundi (Lates calcarifer) DC-SCRIPT, a Specific Molecular Marker for Dendritic Cells in Fish.PLoS One. 2015 Jul 14;10(7):e0132687. doi: 10.1371/journal.pone.0132687. eCollection 2015. PLoS One. 2015. PMID: 26173015 Free PMC article.
-
Programmed cell death of dendritic cells in immune regulation.Immunol Rev. 2010 Jul;236:11-27. doi: 10.1111/j.1600-065X.2010.00916.x. Immunol Rev. 2010. PMID: 20636805 Free PMC article. Review.
Cited by
-
Establishment of Infection Models in Zebrafish Larvae (Danio rerio) to Study the Pathogenesis of Aeromonas hydrophila.Front Microbiol. 2016 Aug 4;7:1219. doi: 10.3389/fmicb.2016.01219. eCollection 2016. Front Microbiol. 2016. PMID: 27540375 Free PMC article.
-
Acute-phase proteins during inflammatory reaction by bacterial infection: Fish-model.Sci Rep. 2019 Mar 18;9(1):4776. doi: 10.1038/s41598-019-41312-z. Sci Rep. 2019. PMID: 30886242 Free PMC article.
-
Studying the immune response to human viral infections using zebrafish.Dev Comp Immunol. 2014 Sep;46(1):84-95. doi: 10.1016/j.dci.2014.03.025. Epub 2014 Apr 6. Dev Comp Immunol. 2014. PMID: 24718256 Free PMC article. Review.
-
Immunogenetic novelty confers a selective advantage in host-pathogen coevolution.Proc Natl Acad Sci U S A. 2018 Feb 13;115(7):1552-1557. doi: 10.1073/pnas.1708597115. Epub 2018 Jan 16. Proc Natl Acad Sci U S A. 2018. PMID: 29339521 Free PMC article.
-
Identification of a Potential Common Ancestor for Mammalian Cross-Presenting Dendritic Cells in Teleost Respiratory Surfaces.Front Immunol. 2018 Jan 25;9:59. doi: 10.3389/fimmu.2018.00059. eCollection 2018. Front Immunol. 2018. PMID: 29422901 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
