

PcrA helicase dismantles RecA filaments by reeling in DNA in uniform steps
- PMID: 20723756
- PMCID: PMC2943210
- DOI: 10.1016/j.cell.2010.07.016
PcrA helicase dismantles RecA filaments by reeling in DNA in uniform steps
Abstract
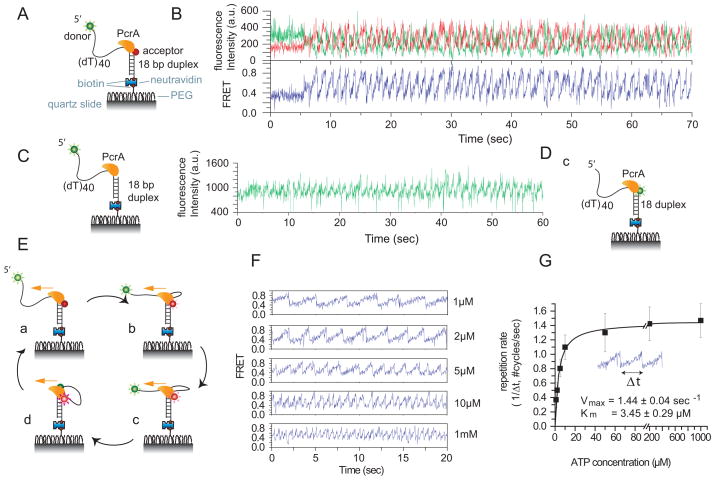
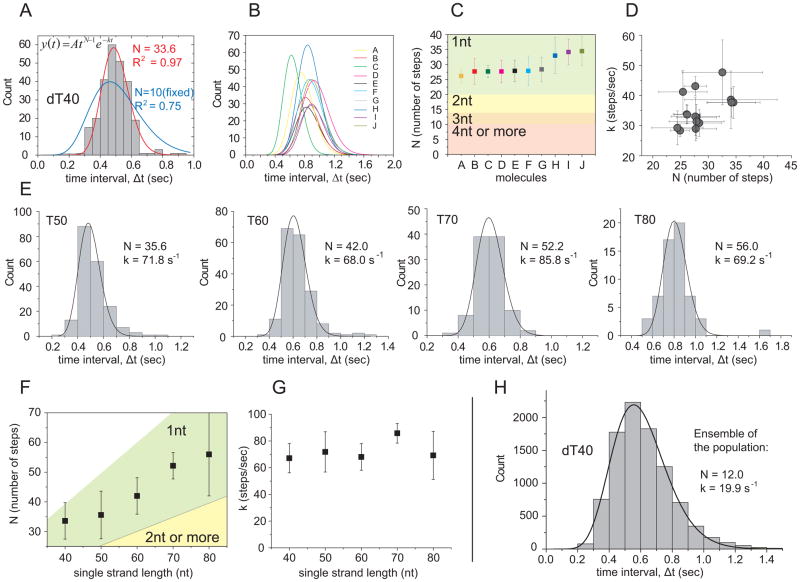
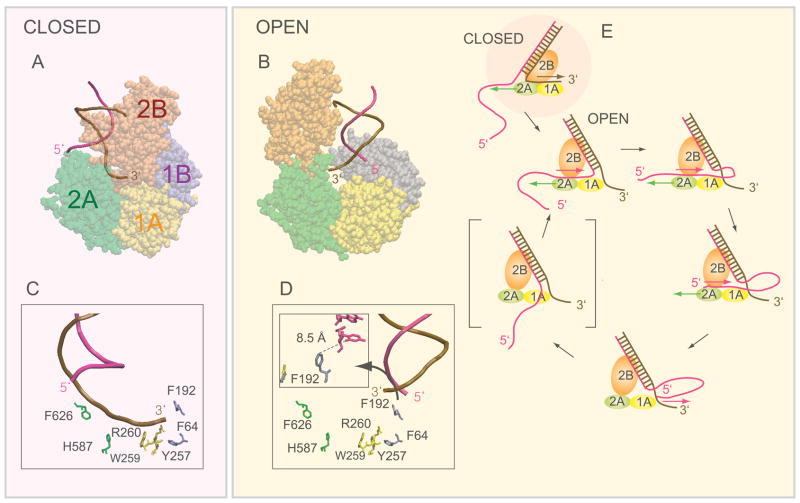
Translocation of helicase-like proteins on nucleic acids underlies key cellular functions. However, it is still unclear how translocation can drive removal of DNA-bound proteins, and basic properties like the elementary step size remain controversial. Using single-molecule fluorescence analysis on a prototypical superfamily 1 helicase, Bacillus stearothermophilus PcrA, we discovered that PcrA preferentially translocates on the DNA lagging strand instead of unwinding the template duplex. PcrA anchors itself to the template duplex using the 2B subdomain and reels in the lagging strand, extruding a single-stranded loop. Static disorder limited previous ensemble studies of a PcrA stepping mechanism. Here, highly repetitive looping revealed that PcrA translocates in uniform steps of 1 nt. This reeling-in activity requires the open conformation of PcrA and can rapidly dismantle a preformed RecA filament even at low PcrA concentrations, suggesting a mode of action for eliminating potentially deleterious recombination intermediates.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
PcrA-mediated disruption of RecA nucleoprotein filaments--essential role of the ATPase activity of RecA.Nucleic Acids Res. 2012 Sep 1;40(17):8416-24. doi: 10.1093/nar/gks641. Epub 2012 Jun 28. Nucleic Acids Res. 2012. PMID: 22743269 Free PMC article.
-
Evidence for a functional dimeric form of the PcrA helicase in DNA unwinding.Nucleic Acids Res. 2008 Apr;36(6):1976-89. doi: 10.1093/nar/gkm1174. Epub 2008 Feb 14. Nucleic Acids Res. 2008. PMID: 18276648 Free PMC article.
-
DNA helicase activity of PcrA is not required for the displacement of RecA protein from DNA or inhibition of RecA-mediated strand exchange.J Bacteriol. 2007 Jun;189(12):4502-9. doi: 10.1128/JB.00376-07. Epub 2007 Apr 20. J Bacteriol. 2007. PMID: 17449621 Free PMC article.
-
Expedient placement of two fluorescent dyes for investigating dynamic DNA protein interactions in real time.Chromosome Res. 2008;16(3):451-67. doi: 10.1007/s10577-008-1235-5. Chromosome Res. 2008. PMID: 18461484 Free PMC article. Review.
-
DNA helicases: 'inching forward'.Curr Opin Struct Biol. 2000 Feb;10(1):124-8. doi: 10.1016/s0959-440x(99)00059-7. Curr Opin Struct Biol. 2000. PMID: 10679457 Review.
Cited by
-
Single-molecule views of protein movement on single-stranded DNA.Annu Rev Biophys. 2012;41:295-319. doi: 10.1146/annurev-biophys-042910-155351. Epub 2012 Feb 23. Annu Rev Biophys. 2012. PMID: 22404684 Free PMC article. Review.
-
Monitoring dynamics of human adenovirus disassembly induced by mechanical fatigue.Sci Rep. 2013;3:1434. doi: 10.1038/srep01434. Sci Rep. 2013. PMID: 23486377 Free PMC article.
-
The Role of SF1 and SF2 Helicases in Biotechnological Applications.Appl Biochem Biotechnol. 2024 Dec;196(12):9064-9084. doi: 10.1007/s12010-024-05027-w. Epub 2024 Aug 2. Appl Biochem Biotechnol. 2024. PMID: 39093351 Review.
-
Monitoring helicase-catalyzed unwinding of multiple duplexes simultaneously.Methods Enzymol. 2022;672:1-27. doi: 10.1016/bs.mie.2022.02.018. Epub 2022 Mar 25. Methods Enzymol. 2022. PMID: 35934470 Free PMC article.
-
Dynamic look at DNA unwinding by a replicative helicase.Proc Natl Acad Sci U S A. 2014 Mar 4;111(9):E827-35. doi: 10.1073/pnas.1322254111. Epub 2014 Feb 18. Proc Natl Acad Sci U S A. 2014. PMID: 24550505 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
