

Analysis of the retinoid isomerase activities in the retinal pigment epithelium and retina
- PMID: 20552438
- PMCID: PMC3049333
- DOI: 10.1007/978-1-60327-325-1_19
Analysis of the retinoid isomerase activities in the retinal pigment epithelium and retina
Abstract
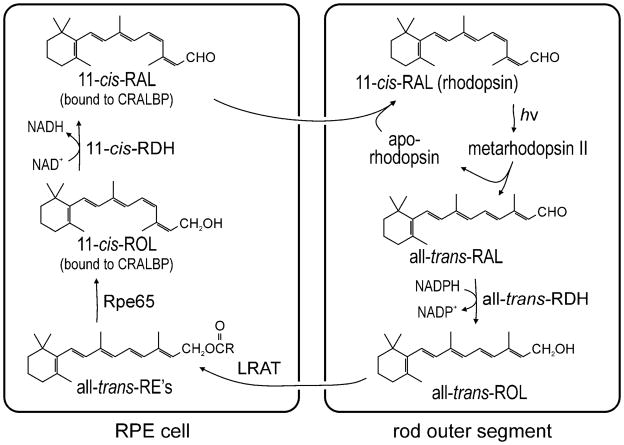
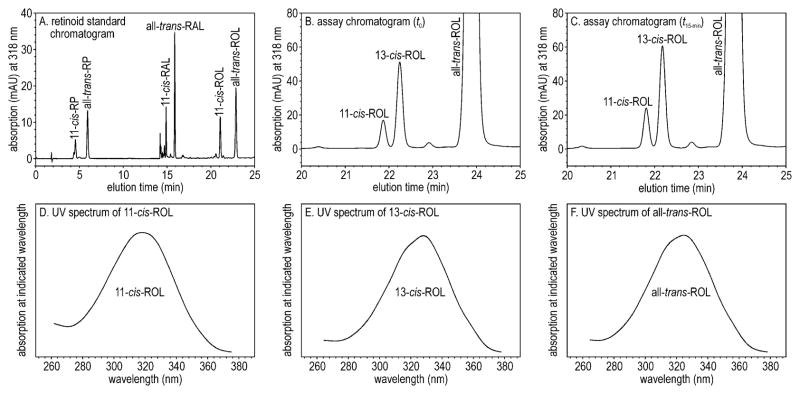
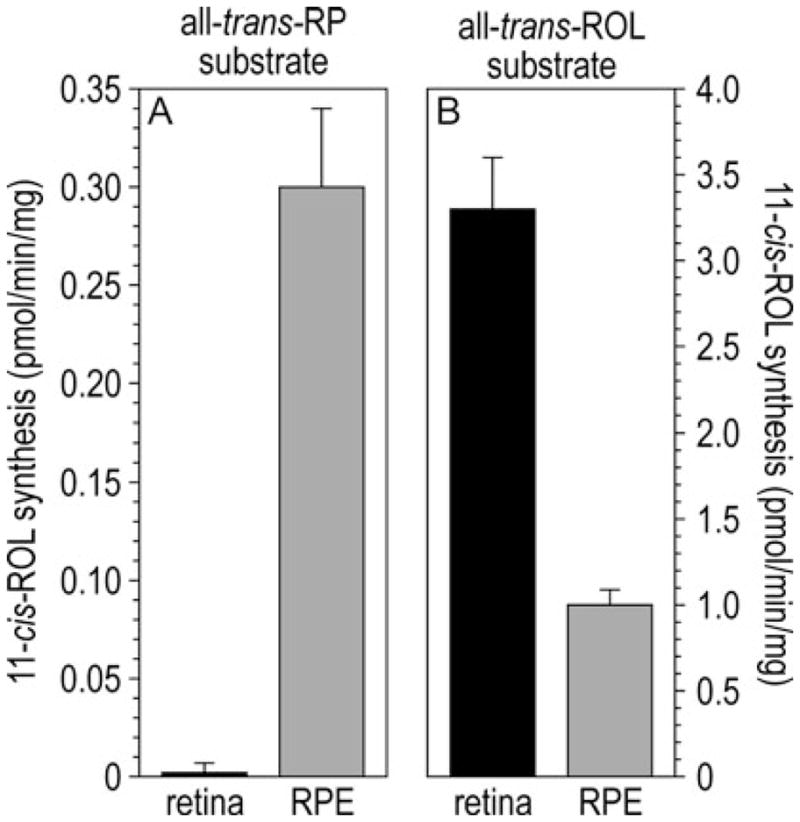
Light sensitivity in the vertebrate retina is mediated by the opsin visual pigments inside rod and cone photoreceptor cells. These pigments consist of a G protein-coupled receptor and the photo-sensitive ligand, 11-cis-retinaldehyde (11-cis-RAL). Absorption of a photon by an opsin pigment induces isomerization of the 11-cis-RAL chromophore to all-trans-retinaldehyde (all-trans-RAL), rendering the pigment insensitive to light. The bleached opsin regains light sensitivity by recombining with another 11-cis-RAL. The vertebrate eye contains a biochemical mechanism for regenerating 11-cis-RAL chromophore from all-trans-RAL, called the visual cycle. The visual cycle takes place within cells of the retinal pigment epithelium (RPE). A second visual cycle also appears to be present in Müller glial cells of the retina. A critical step in the regeneration of 11-cis-RAL chromophore is thermal re-isomerization to the 11-cis configuration of an all-trans-retinyl ester (all-trans-RE) or an all-trans-retinol (all-trans-ROL). In RPE cells, this step is carried out by an enzyme called Rpe65 isomerase. This chapter provides methods for assaying Rpe65 isomerase. Although Rpe65 utilizes an all-trans-RE such as all-trans-retinyl palmitate (all-trans-RP) as substrate, it can be assayed in RPE homogenates by providing all-trans-ROL substrate and allowing the endogenous lecithin:retinol acyl transferase (LRAT) to synthesize all-trans-REs using fatty acids from phosphatidylcholine in the membranes. Alternatively, all-trans-RP can be provided directly as substrate, although this requires the isomerase reaction to be carried out in the presence of detergent, since fatty-acyl esters of all-trans-ROL are insoluble. Methods are provided in this chapter for assaying Rpe65 in RPE homogenates with both all-trans-ROL and all-trans-RP substrates. A second visual cycle appears to be present in the retinas of cone-dominant species such as chicken. This retinal pathway may augment the RPE to provide 11-cis-RAL to cone photoreceptors under conditions of bright light where the rate of opsin photoisomerization is high. The isomerase in this pathway (isomerase-2) utilizes all-trans-ROL and palmitoyl coenzyme A (palm CoA) as substrates to synthesize 11-cis-retinyl palmitate (11-cis-RP). Isomerase-2 appears to be present in Müller cells but has not yet been identified. Methods are provided in this chapter for assaying isomerase-2 in chicken retina homogenates.
Figures
Similar articles
-
Retinal pigment epithelium-retinal G protein receptor-opsin mediates light-dependent translocation of all-trans-retinyl esters for synthesis of visual chromophore in retinal pigment epithelial cells.J Biol Chem. 2008 Jul 11;283(28):19730-8. doi: 10.1074/jbc.M801288200. Epub 2008 May 12. J Biol Chem. 2008. PMID: 18474598 Free PMC article.
-
Identification of the 11-cis-specific retinyl-ester synthase in retinal Müller cells as multifunctional O-acyltransferase (MFAT).Proc Natl Acad Sci U S A. 2014 May 20;111(20):7302-7. doi: 10.1073/pnas.1319142111. Epub 2014 May 5. Proc Natl Acad Sci U S A. 2014. PMID: 24799687 Free PMC article.
-
Chicken retinas contain a retinoid isomerase activity that catalyzes the direct conversion of all-trans-retinol to 11-cis-retinol.Biochemistry. 2005 Sep 6;44(35):11715-21. doi: 10.1021/bi050942m. Biochemistry. 2005. PMID: 16128572 Free PMC article.
-
Vitamin A and Vision.Subcell Biochem. 2016;81:231-259. doi: 10.1007/978-94-024-0945-1_9. Subcell Biochem. 2016. PMID: 27830507 Review.
-
Vitamin A derivatives as treatment options for retinal degenerative diseases.Nutrients. 2013 Jul 12;5(7):2646-66. doi: 10.3390/nu5072646. Nutrients. 2013. PMID: 23857173 Free PMC article. Review.
Cited by
-
Retina, retinol, retinal and the natural history of vitamin A as a light sensor.Nutrients. 2012 Dec 19;4(12):2069-96. doi: 10.3390/nu4122069. Nutrients. 2012. PMID: 23363998 Free PMC article. Review.
-
Visual cycle proteins: Structure, function, and roles in human retinal disease.J Biol Chem. 2018 Aug 24;293(34):13016-13021. doi: 10.1074/jbc.AW118.003228. Epub 2018 Jul 12. J Biol Chem. 2018. PMID: 30002120 Free PMC article.
-
Cultured Müller cells from mammals can synthesize and accumulate retinyl esters.Exp Eye Res. 2012 Aug;101:56-9. doi: 10.1016/j.exer.2012.05.004. Epub 2012 May 24. Exp Eye Res. 2012. PMID: 22634428 Free PMC article. No abstract available.
-
Recent advances in the dark adaptation investigations.Int J Ophthalmol. 2015 Dec 18;8(6):1245-52. doi: 10.3980/j.issn.2222-3959.2015.06.31. eCollection 2015. Int J Ophthalmol. 2015. PMID: 26682182 Free PMC article. Review.
-
The cell biology of the retinal pigment epithelium.Prog Retin Eye Res. 2020 Feb 24:100846. doi: 10.1016/j.preteyeres.2020.100846. Online ahead of print. Prog Retin Eye Res. 2020. PMID: 32105772 Free PMC article. Review.
References
-
- Saari JC, Bredberg DL. Photochemistry and stereoselectivity of cellular retinaldehyde-binding protein from bovine retina. J Biol Chem. 1987;262(16):7618–7622. - PubMed
-
- Eisenfeld AJ, Bunt-Milam AH, Saari JC. Localization of retinoid-binding proteins in developing rat retina. Exp Eye Res. 1985;41(3):299–304. - PubMed
-
- Znoiko SL, et al. Identification of the RPE65 protein in mammalian cone photoreceptors. Invest Ophthal Vis Sci. 2002;43:1604–1609. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
