

Role of the specific interaction of UL112-113 p84 with UL44 DNA polymerase processivity factor in promoting DNA replication of human cytomegalovirus
- PMID: 20538862
- PMCID: PMC2918997
- DOI: 10.1128/JVI.00189-10
Role of the specific interaction of UL112-113 p84 with UL44 DNA polymerase processivity factor in promoting DNA replication of human cytomegalovirus
Abstract
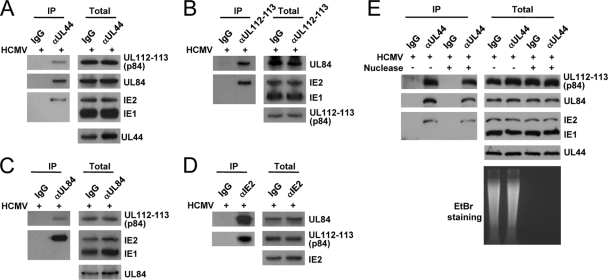
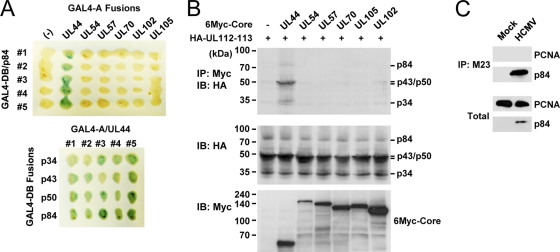
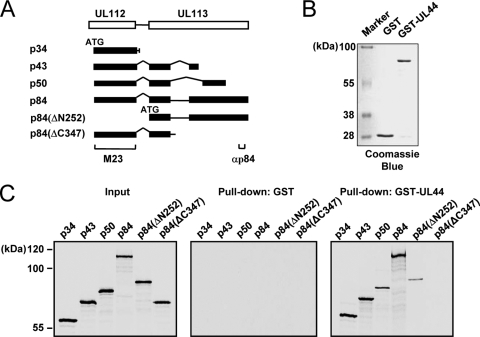
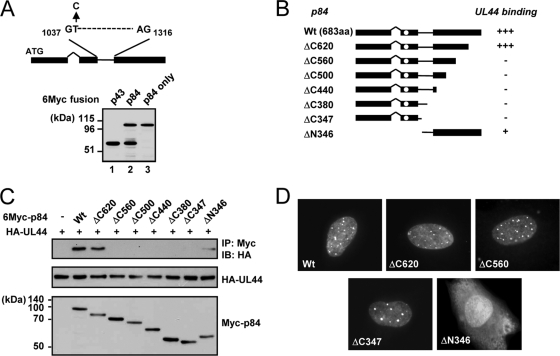
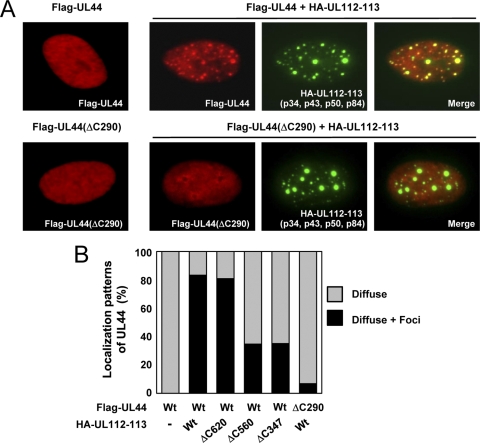
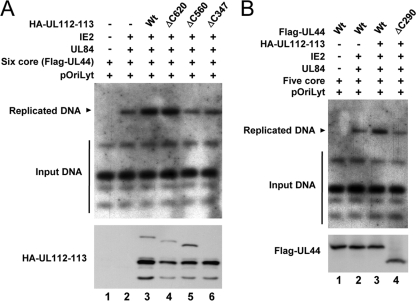
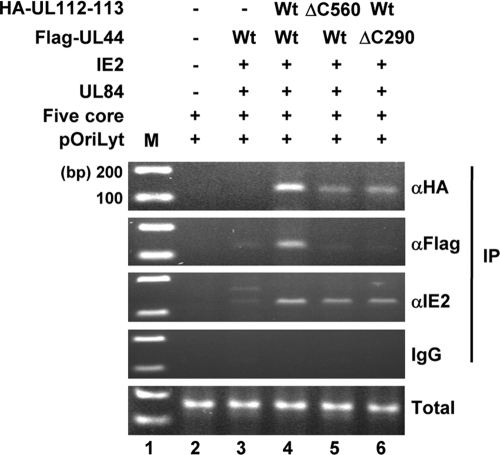
The human cytomegalovirus (HCMV) UL112-113 region encodes four phosphoproteins with common amino termini (p34, p43, p50, and p84) via alternative splicing and is thought to be required for efficient viral DNA replication. We have previously shown that interactions among the four UL112-113 proteins regulate their intranuclear targeting and enable the recruitment of the UL44 DNA polymerase processivity factor to viral prereplication foci. Here, we show that in virus-infected cells, the UL112-113 proteins form a complex with UL44 and other replication proteins, such as UL84 and IE2. In vitro assays showed that all four phosphoproteins interacted with UL44. Interestingly, p84 required both the shared amino-terminal region and the specific near-carboxy-terminal region for UL44 binding. UL44 required both the carboxy-terminal region and the central region, including the dimerization domain for p84 binding. The production of recombinant virus from mutant Towne bacterial artificial chromosome (BAC) DNA, which encodes intact p34, p43, and p50 and a carboxy-terminally truncated p84 defective in UL44 binding, was severely impaired compared to wild-type BAC DNA. A similar defect was observed when mutant BAC DNA encoded a carboxy-terminally truncated UL44 defective in p84 binding. In cotransfection replication assays using six replication core proteins, UL84, IE2, and UL112-113, the efficient replication of an HCMV oriLyt-containing plasmid required the regions of p84 and UL44 necessary for their interaction. Our data suggest that the UL112-113 proteins form a complex with other replication proteins such as UL44, UL84, and IE2 and that the specific interaction of UL112-113 p84 with UL44 is necessary for efficient viral DNA replication.
Figures

Similar articles
-
Differential Requirement of Human Cytomegalovirus UL112-113 Protein Isoforms for Viral Replication.J Virol. 2017 Aug 10;91(17):e00254-17. doi: 10.1128/JVI.00254-17. Print 2017 Sep 1. J Virol. 2017. PMID: 28637762 Free PMC article.
-
Interactions among four proteins encoded by the human cytomegalovirus UL112-113 region regulate their intranuclear targeting and the recruitment of UL44 to prereplication foci.J Virol. 2006 Mar;80(6):2718-27. doi: 10.1128/JVI.80.6.2718-2727.2006. J Virol. 2006. PMID: 16501081 Free PMC article.
-
Requirement of the N-terminal residues of human cytomegalovirus UL112-113 proteins for viral growth and oriLyt-dependent DNA replication.J Microbiol. 2015 Aug;53(8):561-9. doi: 10.1007/s12275-015-5301-3. Epub 2015 Jul 31. J Microbiol. 2015. PMID: 26224459
-
The human cytomegalovirus IE2 and UL112-113 proteins accumulate in viral DNA replication compartments that initiate from the periphery of promyelocytic leukemia protein-associated nuclear bodies (PODs or ND10).J Virol. 1999 Dec;73(12):10458-71. doi: 10.1128/JVI.73.12.10458-10471.1999. J Virol. 1999. PMID: 10559364 Free PMC article.
-
Nuts and bolts of human cytomegalovirus lytic DNA replication.Curr Top Microbiol Immunol. 2008;325:153-66. doi: 10.1007/978-3-540-77349-8_9. Curr Top Microbiol Immunol. 2008. PMID: 18637505 Review.
Cited by
-
Herpes simplex viruses: mechanisms of DNA replication.Cold Spring Harb Perspect Biol. 2012 Sep 1;4(9):a013011. doi: 10.1101/cshperspect.a013011. Cold Spring Harb Perspect Biol. 2012. PMID: 22952399 Free PMC article. Review.
-
Interaction network of proteins associated with human cytomegalovirus IE2-p86 protein during infection: a proteomic analysis.PLoS One. 2013 Dec 16;8(12):e81583. doi: 10.1371/journal.pone.0081583. eCollection 2013. PLoS One. 2013. PMID: 24358118 Free PMC article.
-
A mutation deleting sequences encoding the amino terminus of human cytomegalovirus UL84 impairs interaction with UL44 and capsid localization.J Virol. 2012 Oct;86(20):11066-77. doi: 10.1128/JVI.01379-12. Epub 2012 Aug 1. J Virol. 2012. PMID: 22855486 Free PMC article.
-
Degradation of SAMHD1 Restriction Factor Through Cullin-Ring E3 Ligase Complexes During Human Cytomegalovirus Infection.Front Cell Infect Microbiol. 2020 Jul 30;10:391. doi: 10.3389/fcimb.2020.00391. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32850489 Free PMC article.
-
DNA virus replication compartments.J Virol. 2014 Feb;88(3):1404-20. doi: 10.1128/JVI.02046-13. Epub 2013 Nov 20. J Virol. 2014. PMID: 24257611 Free PMC article. Review.
References
-
- Ahn, J. H., E. J. Brignole III, and G. S. Hayward. 1998. Disruption of PML subnuclear domains by the acidic IE1 protein of human cytomegalovirus is mediated through interaction with PML and may modulate a RING finger-dependent cryptic transactivator function of PML. Mol. Cell. Biol. 18:4899-4913. - PMC - PubMed
-
- Ahn, J. H., and G. S. Hayward. 2000. Disruption of PML-associated nuclear bodies by IE1 correlates with efficient early stages of viral gene expression and DNA replication in human cytomegalovirus infection. Virology 274:39-55. - PubMed
-
- Ahn, J. H., W. J. Jang, and G. S. Hayward. 1999. The human cytomegalovirus IE2 and UL112-113 proteins accumulate in viral DNA replication compartments that initiate from the periphery of promyelocytic leukemia protein-associated nuclear bodies (PODs or ND10). J. Virol. 73:10458-10471. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
