

Interactions between nuclei and the cytoskeleton are mediated by SUN-KASH nuclear-envelope bridges
- PMID: 20507227
- PMCID: PMC4053175
- DOI: 10.1146/annurev-cellbio-100109-104037
Interactions between nuclei and the cytoskeleton are mediated by SUN-KASH nuclear-envelope bridges
Abstract
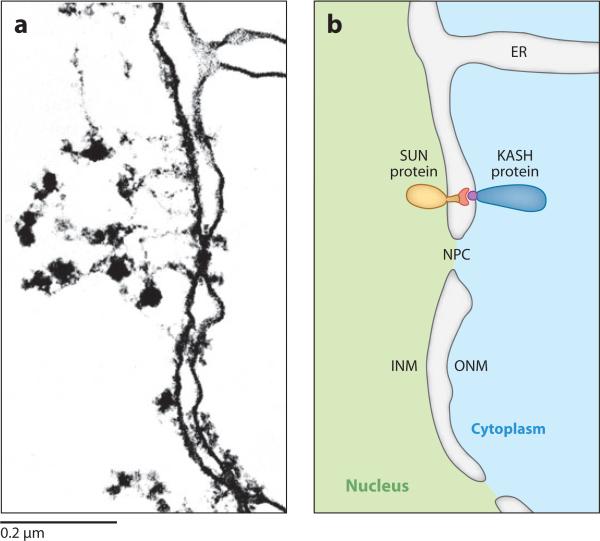
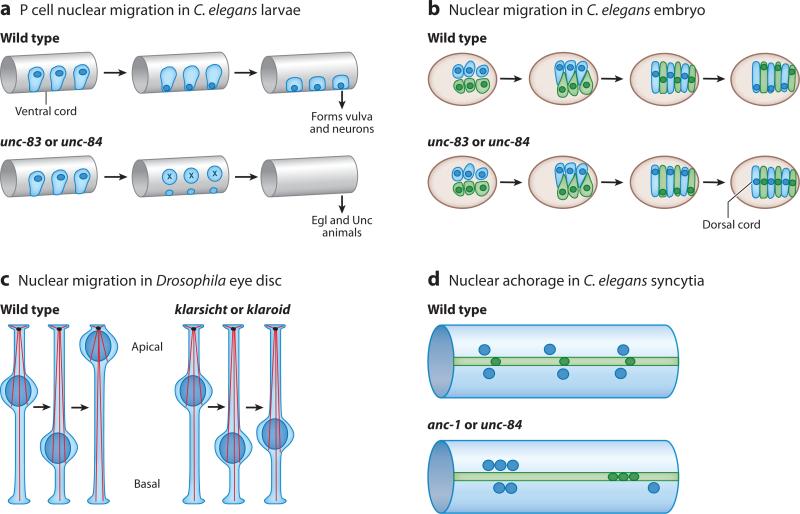
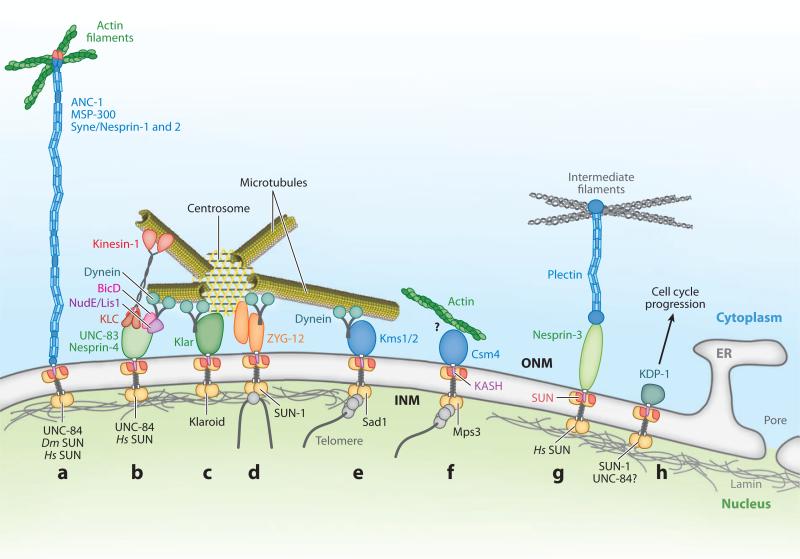
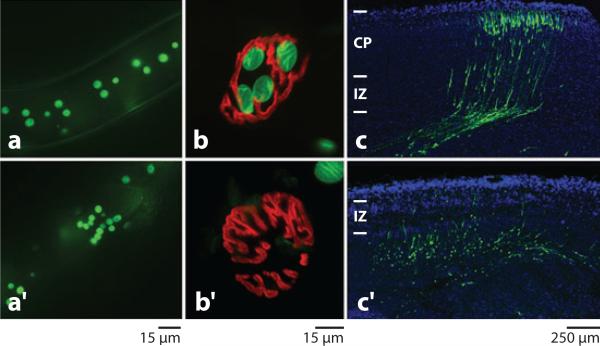
The nuclear envelope links the cytoskeleton to structural components of the nucleus. It functions to coordinate nuclear migration and anchorage, organize chromatin, and aid meiotic chromosome pairing. Forces generated by the cytoskeleton are transferred across the nuclear envelope to the nuclear lamina through a nuclear-envelope bridge consisting of SUN (Sad1 and UNC-84) and KASH (Klarsicht, ANC-1 and Syne/Nesprin homology) proteins. Some KASH-SUN combinations connect microtubules, centrosomes, actin filaments, or intermediate filaments to the surface of the nucleus. Other combinations are used in cell cycle control, nuclear import, or apoptosis. Interactions between the cytoskeleton and the nucleus also affect global cytoskeleton organization. SUN and KASH proteins were identified through genetic screens for mispositioned nuclei in model organisms. Knockouts of SUN or KASH proteins disrupt neurological and muscular development in mice. Defects in SUN and KASH proteins have been linked to human diseases including muscular dystrophy, ataxia, progeria, lissencephaly, and cancer.
Figures
Similar articles
-
Novel plant SUN-KASH bridges are involved in RanGAP anchoring and nuclear shape determination.J Cell Biol. 2012 Jan 23;196(2):203-11. doi: 10.1083/jcb.201108098. J Cell Biol. 2012. PMID: 22270916 Free PMC article.
-
Connecting the nucleus to the cytoskeleton by SUN-KASH bridges across the nuclear envelope.Curr Opin Cell Biol. 2013 Feb;25(1):57-62. doi: 10.1016/j.ceb.2012.10.014. Epub 2012 Nov 10. Curr Opin Cell Biol. 2013. PMID: 23149102 Free PMC article. Review.
-
Conserved SUN-KASH Interfaces Mediate LINC Complex-Dependent Nuclear Movement and Positioning.Curr Biol. 2018 Oct 8;28(19):3086-3097.e4. doi: 10.1016/j.cub.2018.08.001. Epub 2018 Sep 20. Curr Biol. 2018. PMID: 30245107 Free PMC article.
-
UNC-83 IS a KASH protein required for nuclear migration and is recruited to the outer nuclear membrane by a physical interaction with the SUN protein UNC-84.Mol Biol Cell. 2006 Apr;17(4):1790-801. doi: 10.1091/mbc.e05-09-0894. Epub 2006 Feb 15. Mol Biol Cell. 2006. PMID: 16481402 Free PMC article.
-
Communication between the cytoskeleton and the nuclear envelope to position the nucleus.Mol Biosyst. 2007 Sep;3(9):583-9. doi: 10.1039/b703878j. Epub 2007 Jul 16. Mol Biosyst. 2007. PMID: 17700857 Free PMC article. Review.
Cited by
-
High-content imaging with micropatterned multiwell plates reveals influence of cell geometry and cytoskeleton on chromatin dynamics.Biotechnol J. 2015 Oct;10(10):1555-67. doi: 10.1002/biot.201400756. Epub 2015 Jul 14. Biotechnol J. 2015. PMID: 26097126 Free PMC article.
-
Keeping the LINC: the importance of nucleocytoskeletal coupling in intracellular force transmission and cellular function.Biochem Soc Trans. 2011 Dec;39(6):1729-34. doi: 10.1042/BST20110686. Biochem Soc Trans. 2011. PMID: 22103516 Free PMC article. Review.
-
Regulation and coordination of nuclear envelope and nuclear pore complex assembly.Nucleus. 2013 Mar-Apr;4(2):105-14. doi: 10.4161/nucl.23796. Epub 2013 Feb 14. Nucleus. 2013. PMID: 23412657 Free PMC article.
-
Novel plant SUN-KASH bridges are involved in RanGAP anchoring and nuclear shape determination.J Cell Biol. 2012 Jan 23;196(2):203-11. doi: 10.1083/jcb.201108098. J Cell Biol. 2012. PMID: 22270916 Free PMC article.
-
Cytoskeletal interactions at the nuclear envelope mediated by nesprins.Int J Cell Biol. 2012;2012:736524. doi: 10.1155/2012/736524. Epub 2012 Feb 7. Int J Cell Biol. 2012. PMID: 22518138 Free PMC article.
References
-
- Akhtar A, Gasser SM. The nuclear envelope and transcriptional control. Nat. Rev. Genet. 2007;8:507–17. - PubMed
-
- Apel ED, Lewis RM, Grady RM, Sanes JR. Syne-1, a dystrophin- and Klarsicht-related protein associated with synaptic nuclei at the neuromuscular junction. J. Biol. Chem. 2000;275:31986–95. - PubMed
-
- Attali R, Warwar N, Israel A, Gurt I, McNally E, et al. Mutation of SYNE-1, encoding an essential component of the nuclear lamina, is responsible for autosomal recessive arthrogryposis. Hum. Mol. Genet. 2009;18:3462–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
