

Dynamic dependence on ATR and ATM for double-strand break repair in human embryonic stem cells and neural descendants
- PMID: 20368801
- PMCID: PMC2848855
- DOI: 10.1371/journal.pone.0010001
Dynamic dependence on ATR and ATM for double-strand break repair in human embryonic stem cells and neural descendants
Abstract
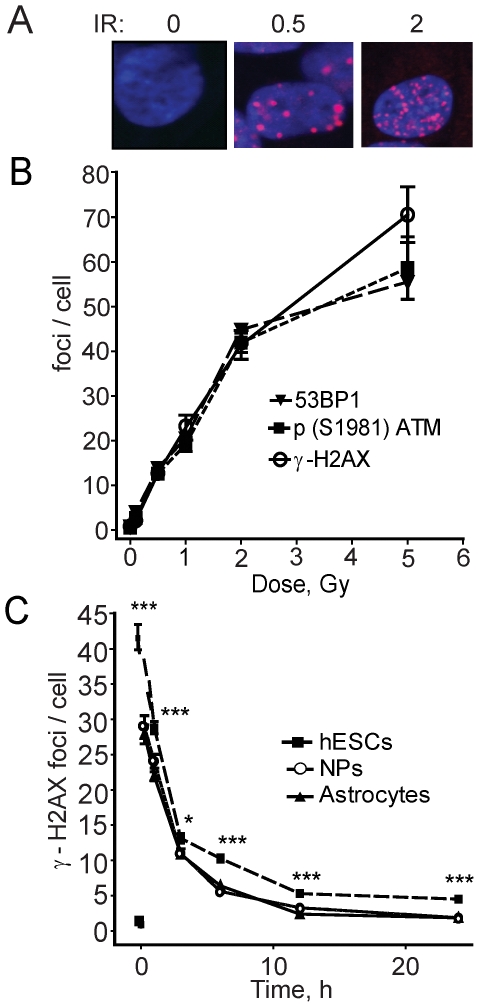
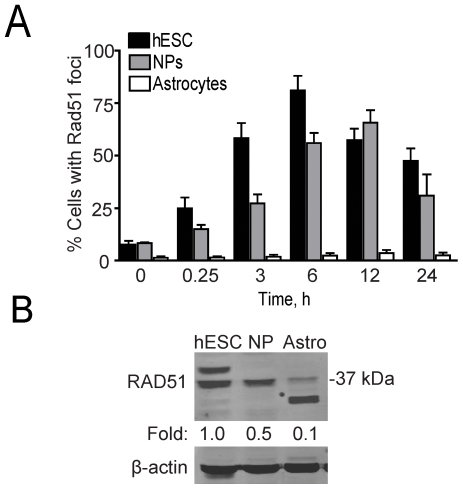
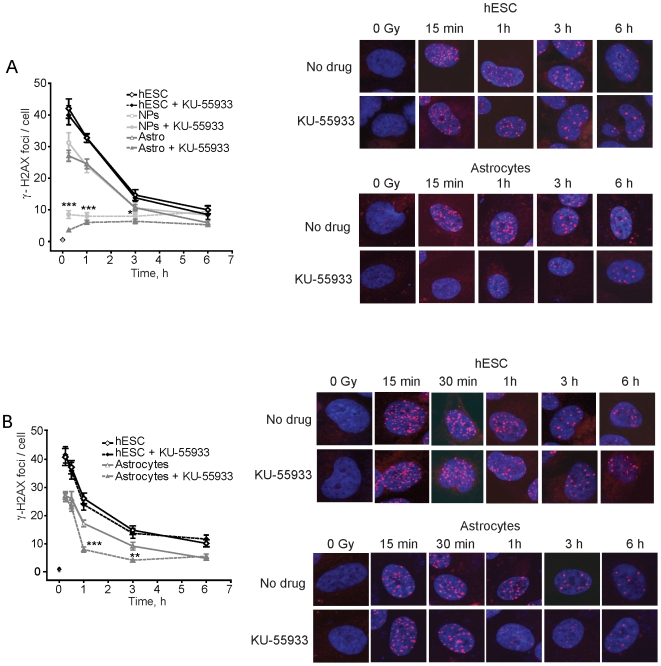
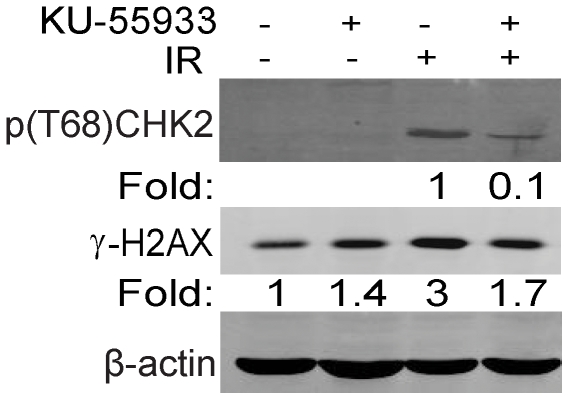
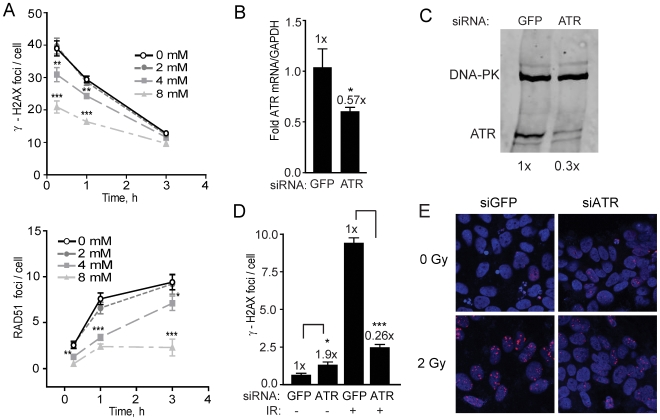
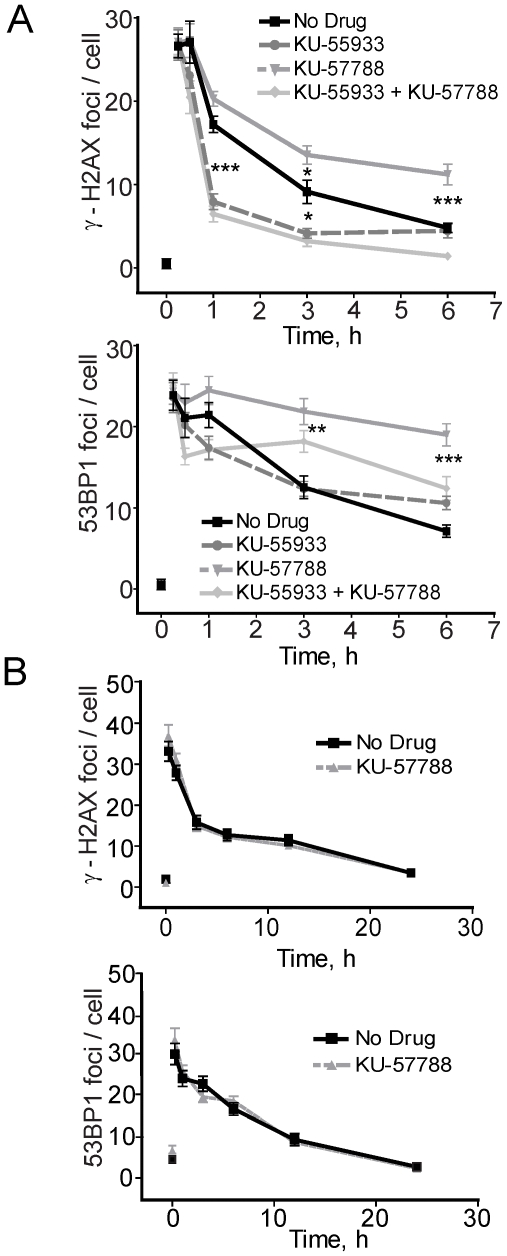
The DNA double-strand break (DSB) is the most toxic form of DNA damage. Studies aimed at characterizing DNA repair during development suggest that homologous recombination repair (HRR) is more critical in pluripotent cells compared to differentiated somatic cells in which nonhomologous end joining (NHEJ) is dominant. We have characterized the DNA damage response (DDR) and quality of DNA double-strand break (DSB) repair in human embryonic stem cells (hESCs), and in vitro-derived neural cells. Resolution of ionizing radiation-induced foci (IRIF) was used as a surrogate for DSB repair. The resolution of gamma-H2AX foci occurred at a slower rate in hESCs compared to neural progenitors (NPs) and astrocytes perhaps reflective of more complex DSB repair in hESCs. In addition, the resolution of RAD51 foci, indicative of active homologous recombination repair (HRR), showed that hESCs as well as NPs have high capacity for HRR, whereas astrocytes do not. Importantly, the ATM kinase was shown to be critical for foci formation in astrocytes, but not in hESCs, suggesting that the DDR is different in these cells. Blocking the ATM kinase in astrocytes not only prevented the formation but also completely disassembled preformed repair foci. The ability of hESCs to form IRIF was abrogated with caffeine and siRNAs targeted against ATR, implicating that hESCs rely on ATR, rather than ATM for regulating DSB repair. This relationship dynamically changed as cells differentiated. Interestingly, while the inhibition of the DNA-PKcs kinase (and presumably non-homologous endjoining [NHEJ]) in astrocytes slowed IRIF resolution it did not in hESCs, suggesting that repair in hESCs does not utilize DNA-PKcs. Altogether, our results show that hESCs have efficient DSB repair that is largely ATR-dependent HRR, whereas astrocytes critically depend on ATM for NHEJ, which, in part, is DNA-PKcs-independent.
Conflict of interest statement
Figures
Similar articles
-
ATM-independent, high-fidelity nonhomologous end joining predominates in human embryonic stem cells.Aging (Albany NY). 2010 Sep;2(9):582-96. doi: 10.18632/aging.100197. Aging (Albany NY). 2010. PMID: 20844317 Free PMC article.
-
DNA-PKcs and ATM co-regulate DNA double-strand break repair.DNA Repair (Amst). 2009 Aug 6;8(8):920-9. doi: 10.1016/j.dnarep.2009.05.006. Epub 2009 Jun 16. DNA Repair (Amst). 2009. PMID: 19535303 Free PMC article.
-
Regulation of ATM in DNA double strand break repair accounts for the radiosensitivity in human cells exposed to high linear energy transfer ionizing radiation.Mutat Res. 2009 Nov 2;670(1-2):15-23. doi: 10.1016/j.mrfmmm.2009.06.016. Epub 2009 Jul 5. Mutat Res. 2009. PMID: 19583974
-
The influence of heterochromatin on DNA double strand break repair: Getting the strong, silent type to relax.DNA Repair (Amst). 2010 Dec 10;9(12):1273-82. doi: 10.1016/j.dnarep.2010.09.013. Epub 2010 Oct 30. DNA Repair (Amst). 2010. PMID: 21036673 Review.
-
Regulation of DNA double-strand break repair pathway choice.Cell Res. 2008 Jan;18(1):134-47. doi: 10.1038/cr.2007.111. Cell Res. 2008. PMID: 18157161 Review.
Cited by
-
Phase I Study of Ceralasertib (AZD6738), a Novel DNA Damage Repair Agent, in Combination with Weekly Paclitaxel in Refractory Cancer.Clin Cancer Res. 2021 Sep 1;27(17):4700-4709. doi: 10.1158/1078-0432.CCR-21-0251. Epub 2021 May 11. Clin Cancer Res. 2021. PMID: 33975862 Free PMC article. Clinical Trial.
-
Achilles' heel of pluripotent stem cells: genetic, genomic and epigenetic variations during prolonged culture.Cell Mol Life Sci. 2016 Jul;73(13):2453-66. doi: 10.1007/s00018-016-2171-8. Epub 2016 Mar 9. Cell Mol Life Sci. 2016. PMID: 26961132 Free PMC article. Review.
-
Human embryonic stem cell responses to ionizing radiation exposures: current state of knowledge and future challenges.Stem Cells Int. 2012;2012:579104. doi: 10.1155/2012/579104. Epub 2012 Aug 16. Stem Cells Int. 2012. PMID: 22966236 Free PMC article.
-
Unraveling the global microRNAome responses to ionizing radiation in human embryonic stem cells.PLoS One. 2012;7(2):e31028. doi: 10.1371/journal.pone.0031028. Epub 2012 Feb 8. PLoS One. 2012. PMID: 22347422 Free PMC article.
-
LINCing Senescence and Nuclear Envelope Changes.Cells. 2022 May 30;11(11):1787. doi: 10.3390/cells11111787. Cells. 2022. PMID: 35681483 Free PMC article. Review.
References
-
- Valerie K, Povirk LF. Regulation and mechanisms of mammalian double-strand break repair. Oncogene. 2003;22:5792–5812. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
