

NuA4-dependent acetylation of nucleosomal histones H4 and H2A directly stimulates incorporation of H2A.Z by the SWR1 complex
- PMID: 20332092
- PMCID: PMC2871465
- DOI: 10.1074/jbc.M110.117069
NuA4-dependent acetylation of nucleosomal histones H4 and H2A directly stimulates incorporation of H2A.Z by the SWR1 complex
Abstract
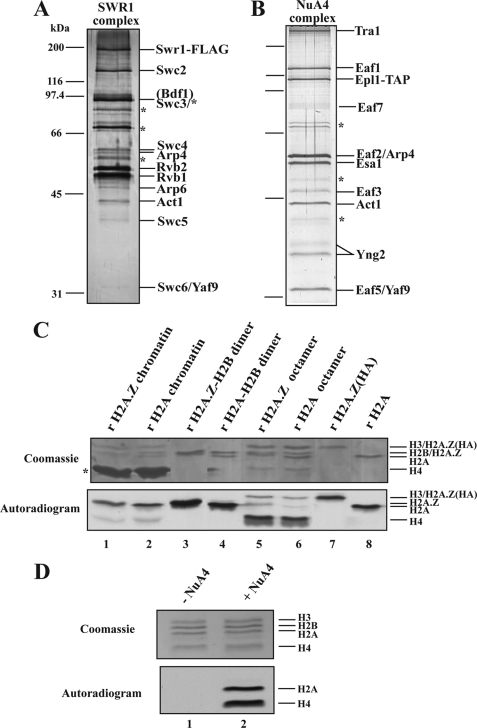
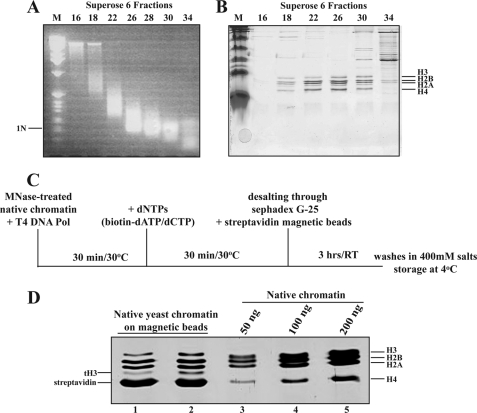
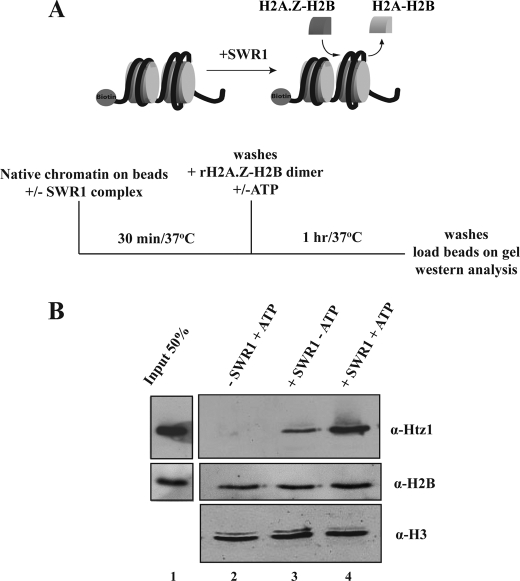
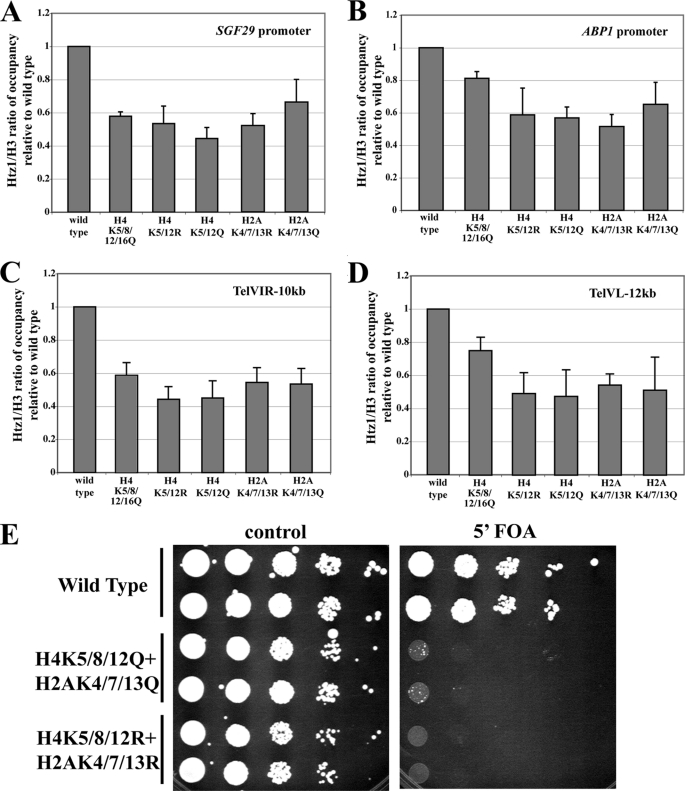
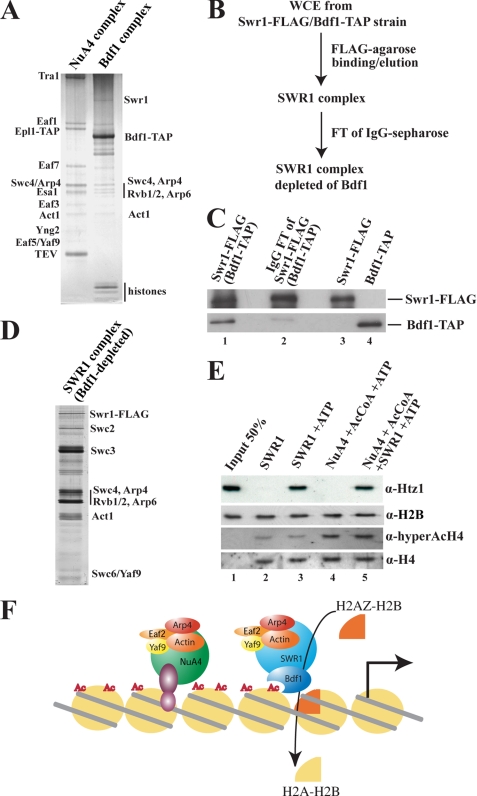
Structural and functional analyses of nucleosomes containing histone variant H2A.Z have drawn a lot of interest over the past few years. Important work in budding yeast has shown that H2A.Z (Htz1)-containing nucleosomes are specifically located on the promoter regions of genes, creating a specific chromatin structure that is poised for disassembly during transcription activation. The SWR1 complex is responsible for incorporation of Htz1 into nucleosomes through ATP-dependent exchange of canonical H2A-H2B dimers for Htz1-H2B dimers. Interestingly, the yeast SWR1 complex is functionally linked to the NuA4 acetyltransferase complex in vivo. NuA4 and SWR1 are physically associated in higher eukaryotes as they are homologous to the TIP60/p400 complex, which encompasses both histone acetyltransferase (Tip60) and histone exchange (p400/Domino) activities. Here we present work investigating the impact of NuA4-dependent acetylation on SWR1-driven incorporation of H2A.Z into chromatin. Using in vitro histone exchange assays with native chromatin, we demonstrate that prior chromatin acetylation by NuA4 greatly stimulates the exchange of H2A for H2A.Z. Interestingly, we find that acetylation of H2A or H4 N-terminal tails by NuA4 can independently stimulate SWR1 activity. Accordingly, we demonstrate that mutations of H4 or H2A N-terminal lysine residues have similar effects on H2A.Z incorporation in vivo, and cells carrying mutations in both tails are nonviable. Finally, depletion experiments indicate that the bromodomain-containing protein Bdf1 is important for NuA4-dependent stimulation of SWR1. These results provide important mechanistic insight into the functional cross-talk between chromatin acetylation and ATP-dependent exchange of histone H2A variants.
Figures


Similar articles
-
Eaf1 is the platform for NuA4 molecular assembly that evolutionarily links chromatin acetylation to ATP-dependent exchange of histone H2A variants.Mol Cell Biol. 2008 Apr;28(7):2257-70. doi: 10.1128/MCB.01755-07. Epub 2008 Jan 22. Mol Cell Biol. 2008. PMID: 18212047 Free PMC article.
-
Drosophila SWR1 and NuA4 complexes are defined by DOMINO isoforms.Elife. 2020 May 20;9:e56325. doi: 10.7554/eLife.56325. Elife. 2020. PMID: 32432549 Free PMC article.
-
Regulation of chromosome stability by the histone H2A variant Htz1, the Swr1 chromatin remodeling complex, and the histone acetyltransferase NuA4.Proc Natl Acad Sci U S A. 2004 Sep 14;101(37):13513-8. doi: 10.1073/pnas.0405753101. Epub 2004 Sep 7. Proc Natl Acad Sci U S A. 2004. PMID: 15353583 Free PMC article.
-
Variation on a theme: Evolutionary strategies for H2A.Z exchange by SWR1-type remodelers.Curr Opin Cell Biol. 2021 Jun;70:1-9. doi: 10.1016/j.ceb.2020.10.014. Epub 2020 Nov 17. Curr Opin Cell Biol. 2021. PMID: 33217681 Review.
-
The specificity of H2A.Z occupancy in the yeast genome and its relationship to transcription.Curr Genet. 2020 Oct;66(5):939-944. doi: 10.1007/s00294-020-01087-7. Epub 2020 Jun 14. Curr Genet. 2020. PMID: 32537667 Review.
Cited by
-
Blocking H2A.Z Incorporation via Tip60 Inhibition Promotes Systems Consolidation of Fear Memory in Mice.eNeuro. 2018 Nov 8;5(5):ENEURO.0378-18.2018. doi: 10.1523/ENEURO.0378-18.2018. eCollection 2018 Sep-Oct. eNeuro. 2018. PMID: 30417078 Free PMC article.
-
Stepwise histone replacement by SWR1 requires dual activation with histone H2A.Z and canonical nucleosome.Cell. 2010 Nov 24;143(5):725-36. doi: 10.1016/j.cell.2010.10.019. Cell. 2010. PMID: 21111233 Free PMC article.
-
Genetic analysis implicates the Set3/Hos2 histone deacetylase in the deposition and remodeling of nucleosomes containing H2A.Z.Genetics. 2011 Apr;187(4):1053-66. doi: 10.1534/genetics.110.125419. Epub 2011 Feb 1. Genetics. 2011. PMID: 21288874 Free PMC article.
-
Eaf5/7/3 form a functionally independent NuA4 submodule linked to RNA polymerase II-coupled nucleosome recycling.EMBO J. 2014 Jun 17;33(12):1397-415. doi: 10.15252/embj.201386433. Epub 2014 May 19. EMBO J. 2014. PMID: 24843044 Free PMC article.
-
Structure and nucleosome interaction of the yeast NuA4 and Piccolo-NuA4 histone acetyltransferase complexes.Nat Struct Mol Biol. 2011 Oct 9;18(11):1196-203. doi: 10.1038/nsmb.2128. Nat Struct Mol Biol. 2011. PMID: 21984211 Free PMC article.
References
-
- Kouzarides T. (2007) Cell 128, 693–705 - PubMed
-
- Clapier C. R., Cairns B. R. (2009) Annu. Rev. Biochem. 78, 273–304 - PubMed
-
- Kamakaka R. T., Biggins S. (2005) Genes Dev. 19, 295–310 - PubMed
-
- Draker R., Cheung P. (2009) Biochem. Cell Biol. 87, 19–25 - PubMed
-
- Raisner R. M., Madhani H. D. (2006) Curr. Opin. Genet. Dev. 16, 119–124 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
