

A novel link between Sus1 and the cytoplasmic mRNA decay machinery suggests a broad role in mRNA metabolism
- PMID: 20230609
- PMCID: PMC2848209
- DOI: 10.1186/1471-2121-11-19
A novel link between Sus1 and the cytoplasmic mRNA decay machinery suggests a broad role in mRNA metabolism
Abstract
Background: Gene expression is achieved by the coordinated action of multiple factors to ensure a perfect synchrony from chromatin epigenetic regulation through to mRNA export. Sus1 is a conserved mRNA export/transcription factor and is a key player in coupling transcription initiation, elongation and mRNA export. In the nucleus, Sus1 is associated to the transcriptional co-activator SAGA and to the NPC associated complex termed TREX2/THSC. Through these associations, Sus1 mediates the nuclear dynamics of different gene loci and facilitate the export of the new transcripts.
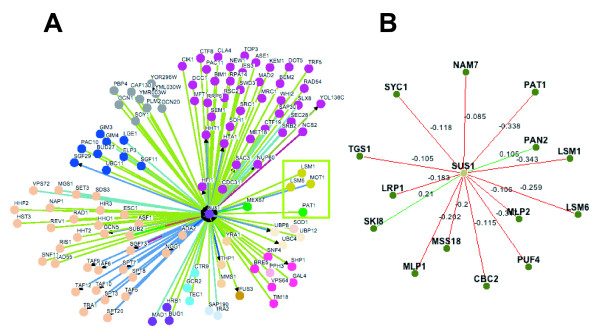
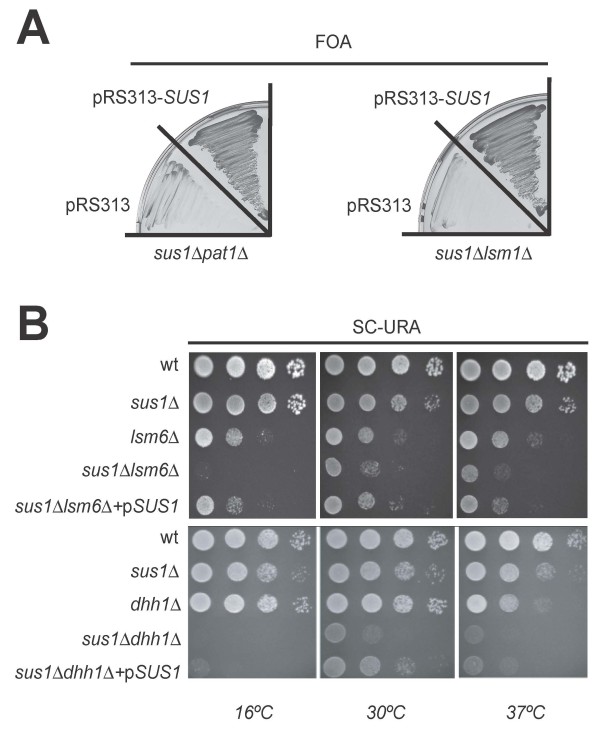
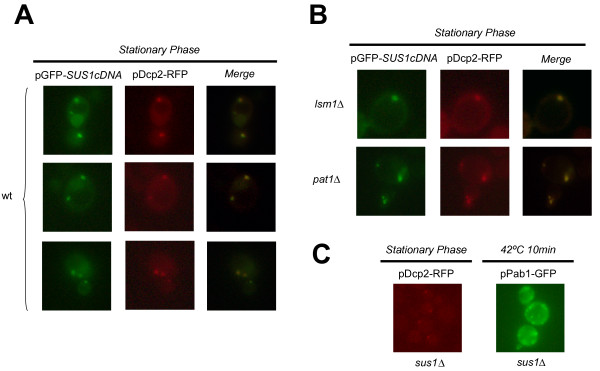
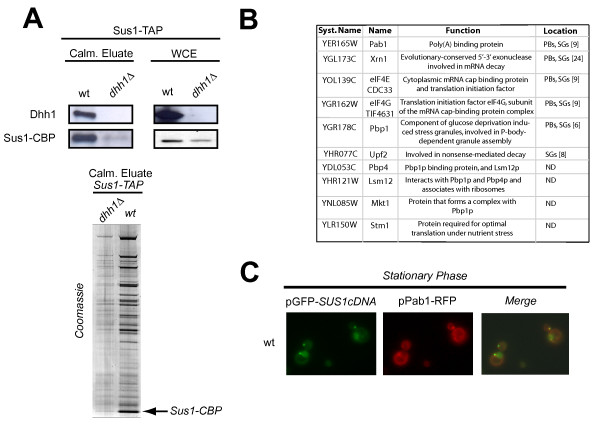
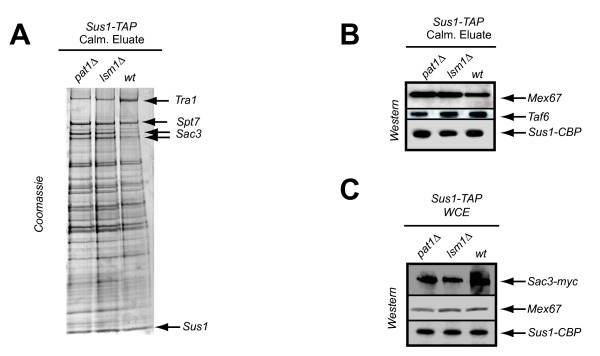
Results: In this study, we have investigated whether the yeast Sus1 protein is linked to factors involved in mRNA degradation pathways. We provide evidence for genetic interactions between SUS1 and genes coding for components of P-bodies such as PAT1, LSM1, LSM6 and DHH1. We demonstrate that SUS1 deletion is synthetic lethal with 5'-->3' decay machinery components LSM1 and PAT1 and has a strong genetic interaction with LSM6 and DHH1. Interestingly, Sus1 overexpression led to an accumulation of Sus1 in cytoplasmic granules, which can co-localise with components of P-bodies and stress granules. In addition, we have identified novel physical interactions between Sus1 and factors associated to P-bodies/stress granules. Finally, absence of LSM1 and PAT1 slightly promotes the Sus1-TREX2 association.
Conclusions: In this study, we found genetic and biochemical association between Sus1 and components responsible for cytoplasmic mRNA metabolism. Moreover, Sus1 accumulates in discrete cytoplasmic granules, which partially co-localise with P-bodies and stress granules under specific conditions. These interactions suggest a role for Sus1 in gene expression during cytoplasmic mRNA metabolism in addition to its nuclear function.
Figures
Similar articles
-
Sus1 is recruited to coding regions and functions during transcription elongation in association with SAGA and TREX2.Genes Dev. 2008 Oct 15;22(20):2811-22. doi: 10.1101/gad.483308. Genes Dev. 2008. PMID: 18923079 Free PMC article.
-
A tale of coupling, Sus1 function in transcription and mRNA export.RNA Biol. 2009 Apr-Jun;6(2):141-4. doi: 10.4161/rna.6.2.7793. Epub 2009 Apr 7. RNA Biol. 2009. PMID: 19246994 Review.
-
Sus1/ENY2: a multitasking protein in eukaryotic gene expression.Crit Rev Biochem Mol Biol. 2012 Nov-Dec;47(6):556-68. doi: 10.3109/10409238.2012.730498. Epub 2012 Oct 12. Crit Rev Biochem Mol Biol. 2012. PMID: 23057668 Review.
-
Genome-wide analysis of RNA-protein interactions illustrates specificity of the mRNA export machinery.Nat Genet. 2003 Feb;33(2):155-61. doi: 10.1038/ng1080. Epub 2003 Jan 13. Nat Genet. 2003. PMID: 12524544
-
Sus1, a functional component of the SAGA histone acetylase complex and the nuclear pore-associated mRNA export machinery.Cell. 2004 Jan 9;116(1):75-86. doi: 10.1016/s0092-8674(03)01025-0. Cell. 2004. PMID: 14718168
Cited by
-
The mRNA degradation factor Xrn1 regulates transcription elongation in parallel to Ccr4.Nucleic Acids Res. 2019 Oct 10;47(18):9524-9541. doi: 10.1093/nar/gkz660. Nucleic Acids Res. 2019. PMID: 31392315 Free PMC article.
-
Predicting the fission yeast protein interaction network.G3 (Bethesda). 2012 Apr;2(4):453-67. doi: 10.1534/g3.111.001560. Epub 2012 Apr 1. G3 (Bethesda). 2012. PMID: 22540037 Free PMC article.
-
A multi-omics dataset of heat-shock response in the yeast RNA binding protein Mip6.Sci Data. 2020 Feb 27;7(1):69. doi: 10.1038/s41597-020-0412-z. Sci Data. 2020. PMID: 32109230 Free PMC article.
-
The evolutionarily conserved factor Sus1/ENY2 plays a role in telomere length maintenance.Curr Genet. 2018 Jun;64(3):635-644. doi: 10.1007/s00294-017-0778-4. Epub 2017 Nov 7. Curr Genet. 2018. PMID: 29116388
-
SUS1 introns are required for efficient mRNA nuclear export in yeast.Nucleic Acids Res. 2011 Oct;39(19):8599-611. doi: 10.1093/nar/gkr496. Epub 2011 Jul 12. Nucleic Acids Res. 2011. PMID: 21749979 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
