

doi: 10.1038/nsmb.1780.
Epub 2010 Mar 14.
Dynamic changes in histone acetylation regulate origins of DNA replication
Affiliations
- PMID: 20228802
- PMCID: PMC3060656
- DOI: 10.1038/nsmb.1780
Item in Clipboard
Dynamic changes in histone acetylation regulate origins of DNA replication
Nat Struct Mol Biol.
2010 Apr.
Abstract
Although histone modifications have been implicated in many DNA-dependent processes, their precise role in DNA replication remains largely unknown. Here we describe an efficient single-step method to specifically purify histones located around an origin of replication from Saccharomyces cerevisiae. Using high-resolution MS, we have obtained a comprehensive view of the histone modifications surrounding the origin of replication throughout the cell cycle. We have discovered that acetylation of histone H3 and H4 is dynamically regulated around an origin of replication, at the level of multiply acetylated histones. Furthermore, we find that this acetylation is required for efficient origin activation during S phase.
Figures
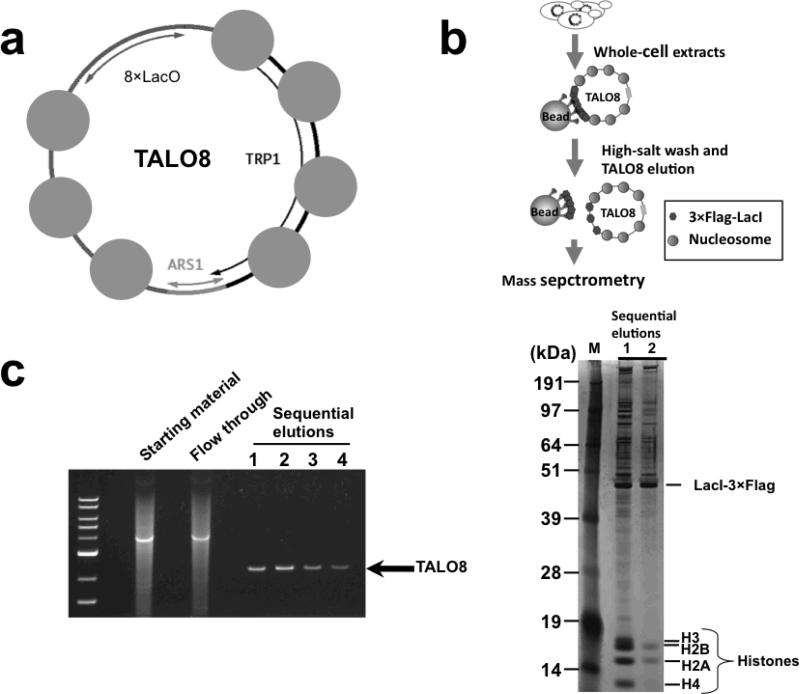
(a) TALO8 is a ~1.8 kb minichromosome with an ARS1 origin of replication, seven well-mapped nucleosomes (depicted as blue circles), a tandem array of eight lac operator sequences (8× Lac O) and a TRP1 selectable marker. (b) A schematic overview of the affinity-purification of TALO8. (c) Representative agarose gel (ethidium bromide-stained) of purified TALO8 DNA linearized by NheI (left) and silver-stained polyacrylamide gel of the purified proteins associated with TALO8 (right). The amount of samples loaded on each elution lane corresponds to 1.5 × 105 and 7.5 × 104 cell equivalents for the agarose and polyacrylamide gels respectively. Typically, eluates 1 and 2 were pooled and prepared for mass spectrometry.
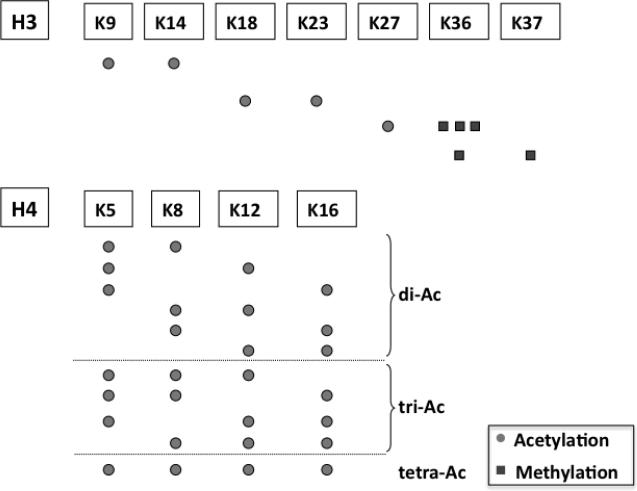
Combinations of histone modifications detected on the same peptide from histones H3 and H4 on TALO8 are shown on the same horizontal row. For lysine methylations, the number of squares indicates the level of methylation.
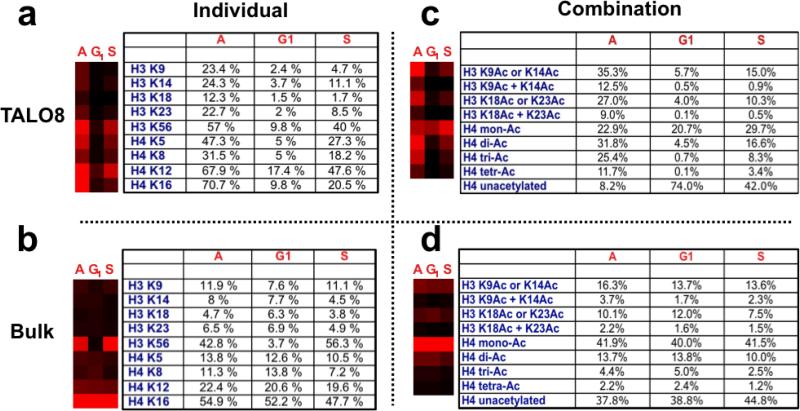
(a) Quantitative analysis of histone acetylations per indicated lysine residue of TALO8-purified histones H3 and H4. Histone acetylation at individual lysines are scored independently of the acetylation states of other lysines. The table indicates the absolute level (in percentage) of histone acetylation detected for each of the indicated residues over the course of the cell cycle. The quantitative data is graphically represented using TreeView, and the intensity of red indicates increasing abundance. “A” refers to asynchronously growing cells. The S phase samples were prepared at 17 minutes after release from α-factor, as done for Figure 2. (b) Quantitative analysis of histone acetylation per lysine residue, as in (a), but on bulk, chromatin-bound histones. (c) Quantitative analysis of combinations of histone acetylations present on TALO8 purified histones. Analysis was done as in (A), with the absolute levels shown in the table and a graphical representation on the left. “H3 K9Ac or K14Ac” indicates peptides containing a single acetyl group which was present either on H3 K9 or H3 K14. Alternatively, “H3 K9Ac K14Ac” indicates di-acetylated peptides where both K9 and K14 were acetylated. The notation is the same for the peptide containing H3 K18 and K23. In the case of histone H4, mono-acetylated amino-terminal peptides (containing K5, K8, K12 and K16) are denoted as “H4 mono Ac”, di-acetylated peptides are “H4 di Ac”, tri-acetylated peptides are “H4 tri Ac” and peptides where K5, K8, K12 and K16 are all acetylated are “H4 tetra Ac”. “A” refers to asynchronously growing cells. The S phase samples were prepared at 17 minutes after release from alpha factor, as done for Figure 2. (d) Quantitative analysis of combinations of histone acetylations, as in (c), but on bulk, chromatin-bound histones.
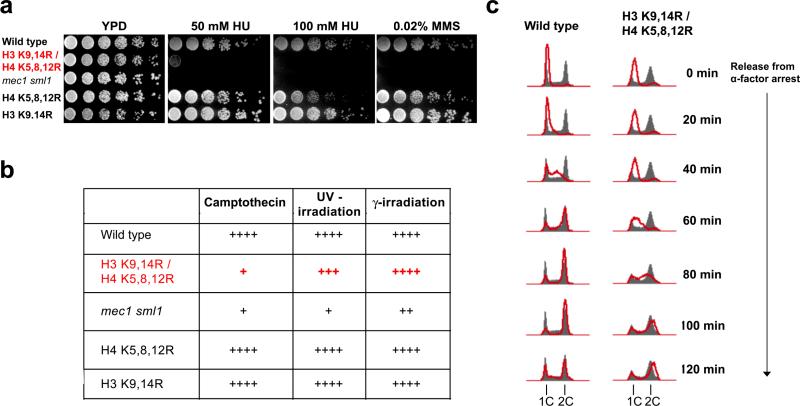
(a) The histone acetylation mutant is hypersensitive to drugs that cause replication stress. Five-fold serial dilutions of yeast strains were spotted onto YPD plates containing 50mM hydroxyurea (HU), 100mM HU, 0.02% methyl methanesulfonate (MMS) or YPD alone and grown at 25°C for 4 days.
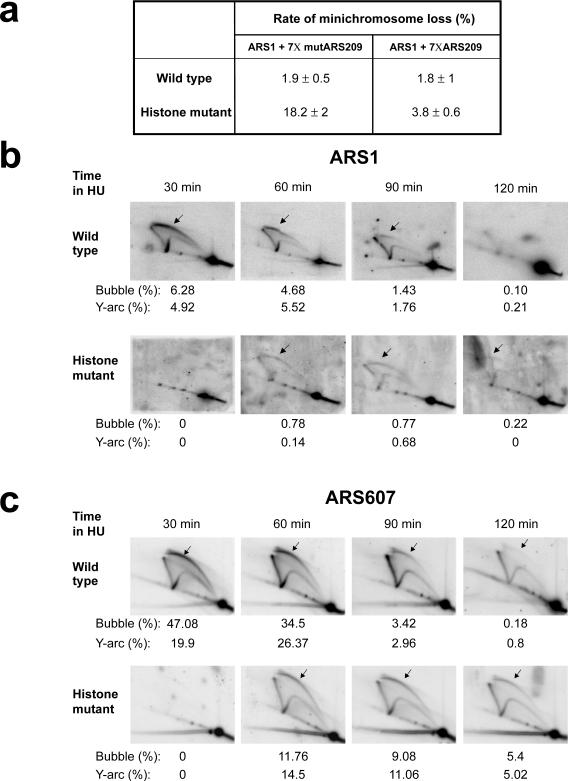
(a) The histone acetylation mutant has an elevated plasmid loss rate. Plasmids containing a functional ARS1 and seven copies of either functional (pDK368-7, right) or mutant ARS209 (pDK398-7, left) were transformed into the histone acetylation mutant and wild type control, and their loss rates were measured as described previously. The numbers denote the mean and the standard deviation of the plasmid loss rates (%) per generation. (b) 2-D agarose gel analyses indicate that the histone mutant is deficient in origin firing. Wild type and histone mutant cells were grown in YPD medium, pre-synchronized in G1 with α-factor at 25°C and released into fresh medium containing 200mM hydroxyurea (HU) and 0.1mg/ml Pronase E. DNA was prepared from cells harvested at the indicated times, digested with NcoI and subjected to electrophoresis in two dimensions. The DNA was transferred onto nylon membranes and probed with a 32P-labelled DNA fragment spanning the ARS1 origin. Quantitation of the abundance of bubble intermediates (indicated by arrows) and Y-arcs as a percentage relative to the 1N spot is given below each image. A schematic representation of relevant replication intermediates is also shown. (c) 2-D agarose gel analyses of another early, efficient origin, ARS607, indicate deficient origin firing in the histone mutant. DNA was digested with BspHI and ClaI and analyzed as in (b), using a 32P-labelled DNA fragment spanning ARS607. Relative quantitation of the replication intermediates (as percentage of the 1N spot intensity) is shown, as in (b).
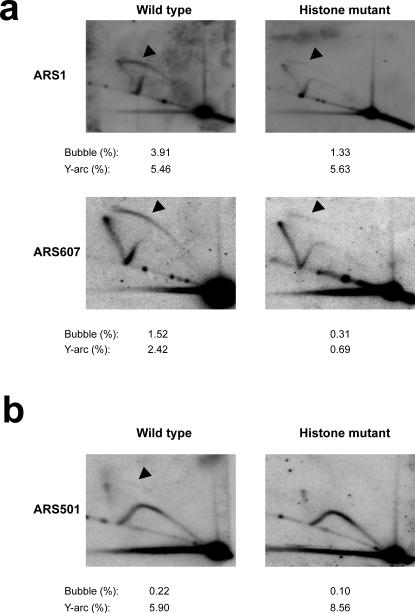
(a) 2-D agarose gel analyses of early origins in the histone mutant under unperturbed growth conditions indicate deficient origin firing. DNA was prepared from pooled S-phase populations of wild type or histone mutant cells, as in Fig 5. NcoI (ARS1) or BspHI ClaI (ARS607) digested DNA was subjected to 2-D gel analyses as before. Relative quantitation of bubble intermediates (represented by arrowheads) and Y-arcs is shown below (as percentage of the 1N spot intensity). (b) Pooled S-phase DNA from (a) was digested with XbaI and subject to 2-D gel analyses. A relative quantitation of bubble intermediate (represented by an arrowhead) and Y-arcs is presented below (as percentage of the 1N spot intensity).
Similar articles
-
Developmentally regulated histone modifications in Drosophila follicle cells: initiation of gene amplification is associated with histone H3 and H4 hyperacetylation and H1 phosphorylation.Chromosoma. 2007 Apr;116(2):197-214. doi: 10.1007/s00412-006-0092-2. Epub 2007 Jan 12. Chromosoma. 2007. PMID: 17219175
-
Control of replication initiation by the Sum1/Rfm1/Hst1 histone deacetylase.BMC Mol Biol. 2008 Nov 6;9:100. doi: 10.1186/1471-2199-9-100. BMC Mol Biol. 2008. PMID: 18990212 Free PMC article.
-
Dynamic changes in chromatin structure through post-translational modifications of histone H3 during replication origin activation.J Cell Biochem. 2009 Oct 1;108(2):400-7. doi: 10.1002/jcb.22266. J Cell Biochem. 2009. PMID: 19585526
-
The activities of eukaryotic replication origins in chromatin.Biochim Biophys Acta. 2004 Mar 15;1677(1-3):142-57. doi: 10.1016/j.bbaexp.2003.11.015. Biochim Biophys Acta. 2004. PMID: 15020055 Review.
-
To fire or not to fire: origin activation in Saccharomyces cerevisiae ribosomal DNA.Genes Dev. 2002 Oct 1;16(19):2459-64. doi: 10.1101/gad.1033702. Genes Dev. 2002. PMID: 12368256 Review. No abstract available.
Cited by
-
Proteomics in epigenetics: new perspectives for cancer research.Brief Funct Genomics. 2013 May;12(3):205-18. doi: 10.1093/bfgp/elt002. Epub 2013 Feb 11. Brief Funct Genomics. 2013. PMID: 23401080 Free PMC article. Review.
-
Methylation of histone H3 at lysine 37 by Set1 and Set2 prevents spurious DNA replication.Mol Cell. 2021 Jul 1;81(13):2793-2807.e8. doi: 10.1016/j.molcel.2021.04.021. Epub 2021 May 11. Mol Cell. 2021. PMID: 33979575 Free PMC article.
-
Increased transcription in hydroxyurea-treated root meristem cells of Vicia faba.Protoplasma. 2013 Feb;250(1):251-9. doi: 10.1007/s00709-012-0402-x. Epub 2012 Apr 15. Protoplasma. 2013. PMID: 22526201 Free PMC article.
-
Exploiting Replication Stress as a Novel Therapeutic Intervention.Mol Cancer Res. 2021 Feb;19(2):192-206. doi: 10.1158/1541-7786.MCR-20-0651. Epub 2020 Oct 5. Mol Cancer Res. 2021. PMID: 33020173 Free PMC article. Review.
-
Pervasive transcription fine-tunes replication origin activity.Elife. 2018 Dec 17;7:e40802. doi: 10.7554/eLife.40802. Elife. 2018. PMID: 30556807 Free PMC article.
References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–60. see comments. - PubMed
-
- Groth A, Rocha W, Verreault A, Almouzni G. Chromatin challenges during DNA replication and repair. Cell. 2007;128:721–33. - PubMed
-
- Li B, Carey M, Workman JL. The role of chromatin during transcription. Cell. 2007;128:707–19. - PubMed
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- Stinchcomb DT, Struhl K, Davis RW. Isolation and characterisation of a yeast chromosomal replicator. Nature. 1979;282:39–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
