

Review
doi: 10.1242/dev.039487.
Maternal control of early mouse development
Affiliations
- PMID: 20179092
- PMCID: PMC2834456
- DOI: 10.1242/dev.039487
Item in Clipboard
Review
Maternal control of early mouse development
Development.
2010 Mar.
Abstract
The hiatus between oocyte and embryonic gene transcription dictates a role for stored maternal factors in early mammalian development. Encoded by maternal-effect genes, these factors accumulate during oogenesis and enable the activation of the embryonic genome, the subsequent cleavage stages of embryogenesis and the initial establishment of embryonic cell lineages. Recent studies in mice have yielded new findings on the role of maternally provided proteins and multi-component complexes in preimplantation development. Nevertheless, significant gaps remain in our mechanistic understanding of the networks that regulate early mammalian embryogenesis, which provide an impetus and opportunities for future investigations.
Figures
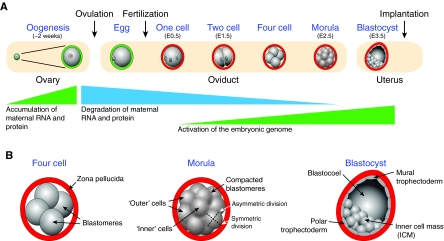
Preimplantation mouse development. (A) Oogenesis and early development. Maternal RNA and proteins accumulate within individual oocytes during a ~2 week growth phase, but many are degraded during meiotic maturation and ovulation. The zona pellucida surrounds growing oocytes and ovulated eggs (green) and is modified following fertilization (red) to prevent polyspermy and to protect the embryo as it passes through the oviduct. At embryonic day 0.5 (E0.5), 1-cell embryos can be recovered from the oviduct and, by the 2-cell stage (E1.5), there is robust embryonic genome activation. Following the third cell division, embryos undergo a Ca2+-mediated compaction to form morulae (E2.5), and develop a fluid-filled blastocoel cavity from the 32-cell stage to become blastocysts (E3.5) that implant in the uterus wall (E4.5). (B) Late preimplantation development. At four cells, blastomeres with distinct cell boundaries are encased within the protective confines of the zona pellucida. Individual cells remain pluripotent. Following the 8-cell stage, the embryo compacts to form the morula, with increased cell contact that morphologically obscures cell boundaries. Symmetric cell divisions result in similar daughter cells. Asymmetric divisions provide topologically distinct outer cells, which are trophectoderm precursors (forming the placenta), and inner cells, which establish the inner cell mass (ICM) of the fetus. Blastocoel formation defines the blastocyst, and the eccentric localization of the ICM localizes the polar and mural trophectoderm, the latter of which initiates uterine implantation.
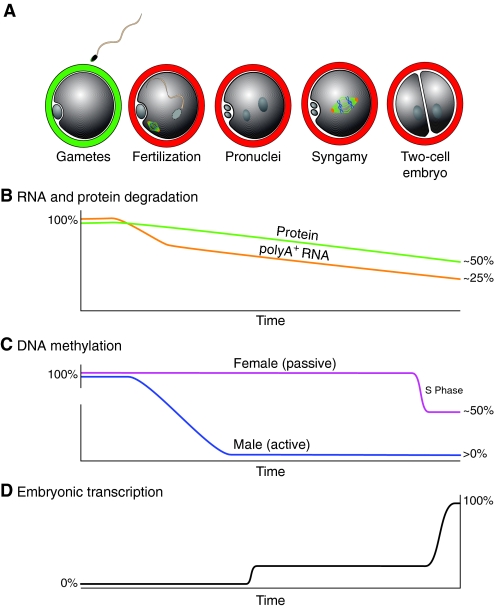
Molecular changes initiate development. (A) The onset of mouse development. Transcriptionally inert gametes fuse at fertilization and the entire sperm is incorporated into the cytoplasm. The sperm nucleus is decondensed and repackaged with egg-stored histones to form the male pronucleus. The egg completes its second meiotic division and forms the female pronuclei. After syngamy and chromosome congression, cytokinesis establishes the 2-cell embryo. (B) RNA and protein degradation. Maternal stores of RNA and protein are required for cleavage-stage development, but these stores are degraded over time, leaving ~25% of maternal polyA+ RNA and ~50% of maternal proteins in the 2-cell embryo (Merz et al., 1981; Clegg and Piko, 1983; De Leon et al., 1983). (C) Demethylation of the embryonic genome. After fertilization, the male genome is actively demethylated prior to DNA replication and male pronucleus formation. By contrast, the female genome undergoes passive demethylation with each cell division (beginning with the first), resulting in cleavage-stage embryos with hypomethylated DNA, except at imprinting control regions (see Glossary, Box 1) (Reik et al., 2001). (D) Embryonic gene activation. Following a minor activation of transcription at the pronuclear stage (more in the male than the female pronucleus), embryonic transcription is robustly activated at the 2-cell stage.

Proteins of the subcortical maternal complex (SCMC). (A) MATER is the largest protein in the complex and has a novel 5× N-terminal repeat, a NACHT domain (http://pfam.sanger.ac.uk/ ) and a 13× leucine-rich repeat that is implicated in protein-protein interactions. (B) FLOPED is the smallest SCMC protein and contains a single, atypical KH domain. (C) PADI6 is an enzyme that converts arginine to citrulline; its specific substrates have yet to be identified in the early embryo. (D) TLE6 is a member of the Groucho co-repressor family that is unique to mammals and lacks the N-terminal Q domain found in other members. NLS, nuclear localization signal. (E) FILIA binds to MATER, but not to FLOPED or TLE6. For a summary of null phenotypes, see Table 2. These proteins are encoded by: Filia, official symbol 2410004A20Rik; Floped, factor located in oocytes permitting embryonic development, official symbol Ooep; Mater, maternal antigen that embryos require, official symbol Nlrp5; Padi6, peptidylarginine deiminase 6; Tle6, transducin-like enhancer of split 6.
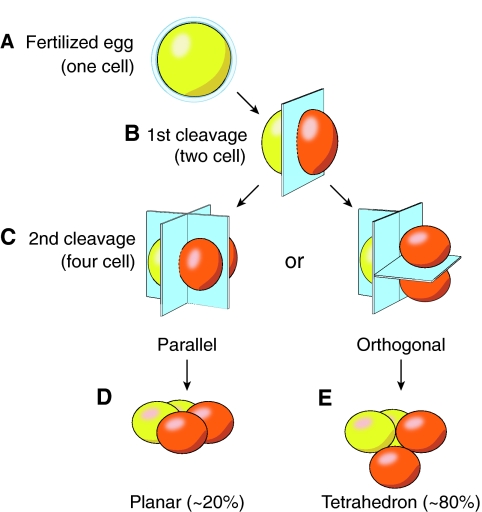
Initial embryonic cleavages. (A,B) The first cleavage of the fertilized egg forms the symmetric 2-cell embryo, in which each blastomere (orange and red) remains totipotent. (C) The second embryonic cleavage occurs sequentially and can either be parallel to the first cleavage (left) or orthogonal to the first cleavage (right). (D) If the former occurs, the pluripotent blastomeres of the 4-cell embryo are present on a single plane. (E) Most commonly (~80%), however, orthogonal division occurs and the blastomeres form a tetrahedron.
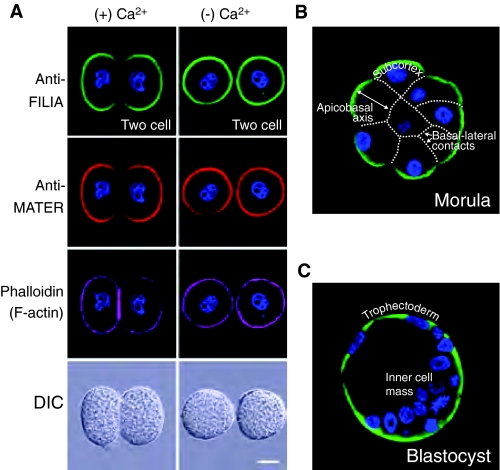
Localization of the SCMC in the early mouse embryo. (A) Exclusion from cell-cell contacts. Zona-pellucida-free 2-cell mouse embryos incubated in the presence (+) or absence (−) of calcium, and fixed, permeabilized and stained with antibodies to FILIA (green), MATER (red) or with phalloidin to stain F-actin (purple), prior to imaging by confocal microscopy or differential interference optics (DIC). The exclusion of the SCMC from regions of cell-cell contact of 2-cell embryos (left) is readily reversible upon disaggregation of the two blastomeres in the absence of calcium (right). Scale bar: 20 μm. (B) Absence of SCMC from inner cells. Morula fixed, permeabilized and stained with antibody to FILIA prior to imaging by confocal microscopy. Dotted lines superimposed on the morula represent basal-lateral contacts of cell membranes. The SCMC (as defined by antibodies to FILIA) is located in the subcortex of outer cells that are polarized (apicobasal) and is not detected in the non-polarized inner cells of the morula. (C) Same as B, but showing a blastocyst. The restriction of the SCMC to the outer cells of the blastocyst provides a marker for trophectoderm cells, which give rise to the placenta. The SCMC is excluded from the ICM. Modified, with permission, from Ohsugi et al. (Ohsugi et al., 2008).
Similar articles
-
[Crucial role of Polycomb proteins from maternal origin in mouse early embryonic development].Med Sci (Paris). 2012 Dec;28(12):1047-9. doi: 10.1051/medsci/20122812009. Epub 2012 Dec 21. Med Sci (Paris). 2012. PMID: 23290399 French. No abstract available.
-
What is the role of maternally provided Cdx2 mRNA in early mouse embryogenesis?Reprod Biomed Online. 2011 Jun;22(6):512-5. doi: 10.1016/j.rbmo.2011.02.003. Epub 2011 Feb 13. Reprod Biomed Online. 2011. PMID: 21498124
-
Post-translational regulation of the maternal-to-zygotic transition.Cell Mol Life Sci. 2018 May;75(10):1707-1722. doi: 10.1007/s00018-018-2750-y. Epub 2018 Feb 9. Cell Mol Life Sci. 2018. PMID: 29427077 Free PMC article. Review.
-
Degradation and translation of maternal mRNA for embryogenesis.Trends Genet. 2024 Mar;40(3):238-249. doi: 10.1016/j.tig.2023.12.008. Epub 2024 Jan 22. Trends Genet. 2024. PMID: 38262796 Review.
-
Molecular determinants of oocyte competence: potential functional role for maternal (oocyte-derived) follistatin in promoting bovine early embryogenesis.Endocrinology. 2009 May;150(5):2463-71. doi: 10.1210/en.2008-1574. Epub 2009 Jan 29. Endocrinology. 2009. PMID: 19179440
Cited by
-
The Maternal Effect Genes UTX and JMJD3 Play Contrasting Roles in Mus musculus Preimplantation Embryo Development.Sci Rep. 2016 Jul 7;6:26711. doi: 10.1038/srep26711. Sci Rep. 2016. PMID: 27384759 Free PMC article.
-
Low-input lipidomics reveals lipid metabolism remodelling during early mammalian embryo development.Nat Cell Biol. 2024 Feb;26(2):278-293. doi: 10.1038/s41556-023-01341-3. Epub 2024 Feb 1. Nat Cell Biol. 2024. PMID: 38302721
-
Preliminary crystallographic analysis of the N-terminal domain of FILIA, a protein essential for embryogenesis.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010 Sep 1;66(Pt 9):1111-4. doi: 10.1107/S1744309110031994. Epub 2010 Aug 28. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010. PMID: 20823540 Free PMC article.
-
LSM14B controls oocyte mRNA storage and stability to ensure female fertility.Cell Mol Life Sci. 2023 Aug 14;80(9):247. doi: 10.1007/s00018-023-04898-2. Cell Mol Life Sci. 2023. PMID: 37578641 Free PMC article.
-
Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis).Int J Mol Sci. 2022 Aug 3;23(15):8605. doi: 10.3390/ijms23158605. Int J Mol Sci. 2022. PMID: 35955739 Free PMC article.
References
-
- Adenot P. G., Mercier Y., Renard J. P., Thompson E. M. (1997). Differential H4 acetylation of paternal and maternal chromatin precedes DNA replication and differential transcriptional activity in pronuclei of 1-cell mouse embryos. aDevelopment 124, 4615-4625 - PubMed
-
- Alizadeh Z., Kageyama S., Aoki F. (2005). Degradation of maternal mRNA in mouse embryos: selective degradation of specific mRNAs after fertilization. Mol. Reprod. Dev. 72, 281-290 - PubMed
-
- Barton S. C., Surani M. A., Norris M. L. (1984). Role of paternal and maternal genomes in mouse development. Nature 311, 374-376 - PubMed
-
- Berryman M., Franck Z., Bretscher A. (1993). Ezrin is concentrated in the apical microvilli of a wide variety of epithelial cells whereas moesin is found primarily in endothelial cells. J. Cell Sci. 105, 1025-1043 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
