

Role of the GTPase Rab1b in ebolavirus particle formation
- PMID: 20164217
- PMCID: PMC2863720
- DOI: 10.1128/JVI.00010-10
Role of the GTPase Rab1b in ebolavirus particle formation
Abstract
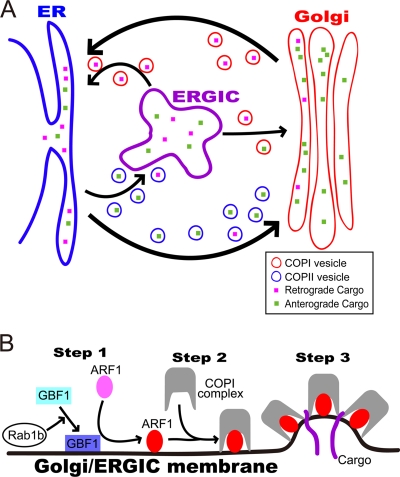
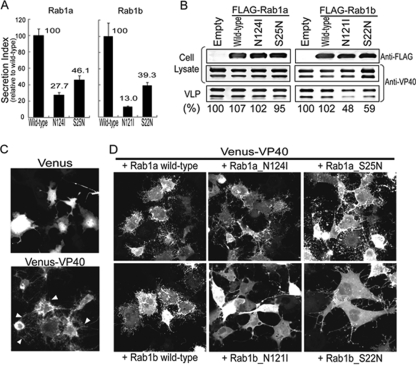
The Ebolavirus matrix protein VP40 is essential for virion assembly and egress. Recently, we reported that the coat protein complex II (COPII) transport system plays an important role in the transport of VP40 to the plasma membrane. Here, we show that dominant-negative mutants of the GTPase Rab1b interfere with VP40-mediated particle formation. Rab1b activates GBF1 (Golgi-specific BFA [brefeldin A] resistance factor 1), a critical factor in the assembly of COPI vesicles. Activated GBF1 stimulates ARF1 (ADP ribosylation factor 1), which recruits coat protein to cellular membranes for the assembly of COPI vesicles. Here, we demonstrate that GBF1 and ARF1 are involved in Ebolavirus virion formation, suggesting that both the COPII and COPI transport systems play a role in Ebolavirus VP40-mediated particle formation. These findings provide new insights into the cellular pathways employed for Ebolavirus virion formation.
Figures
Similar articles
-
Rab1b interacts with GBF1 and modulates both ARF1 dynamics and COPI association.Mol Biol Cell. 2007 Jul;18(7):2400-10. doi: 10.1091/mbc.e06-11-1005. Epub 2007 Apr 11. Mol Biol Cell. 2007. PMID: 17429068 Free PMC article.
-
Rab1b-GBF1-ARF1 Secretory Pathway Axis Is Required for Birnavirus Replication.J Virol. 2022 Feb 23;96(4):e0200521. doi: 10.1128/JVI.02005-21. Epub 2021 Dec 8. J Virol. 2022. PMID: 34878889 Free PMC article.
-
COPI recruitment is modulated by a Rab1b-dependent mechanism.Mol Biol Cell. 2003 May;14(5):2116-27. doi: 10.1091/mbc.e02-09-0625. Epub 2003 Feb 6. Mol Biol Cell. 2003. PMID: 12802079 Free PMC article.
-
GBF1 and Arf1 function in vesicular trafficking, lipid homoeostasis and organelle dynamics.Biol Cell. 2017 Dec;109(12):391-399. doi: 10.1111/boc.201700042. Epub 2017 Nov 6. Biol Cell. 2017. PMID: 28985001 Review.
-
Membrane curvature and the control of GTP hydrolysis in Arf1 during COPI vesicle formation.Biochem Soc Trans. 2005 Aug;33(Pt 4):619-22. doi: 10.1042/BST0330619. Biochem Soc Trans. 2005. PMID: 16042557 Review.
Cited by
-
Viperin Targets Flavivirus Virulence by Inducing Assembly of Noninfectious Capsid Particles.J Virol. 2017 Dec 14;92(1):e01751-17. doi: 10.1128/JVI.01751-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29046456 Free PMC article.
-
Tick-Borne Flaviviruses and the Type I Interferon Response.Viruses. 2018 Jun 21;10(7):340. doi: 10.3390/v10070340. Viruses. 2018. PMID: 29933625 Free PMC article. Review.
-
Role of Coatomer Protein I in Virus Replication.J Virol Antivir Res. 2012 Oct 30;1(2):10.4172/2324-8955.1000102. doi: 10.4172/2324-8955.1000102. J Virol Antivir Res. 2012. PMID: 24392458 Free PMC article.
-
Influenza infection modulates vesicular trafficking and induces Golgi complex disruption.Virusdisease. 2016 Dec;27(4):357-368. doi: 10.1007/s13337-016-0347-3. Epub 2016 Sep 14. Virusdisease. 2016. PMID: 28004015 Free PMC article.
-
Viral protein engagement of GBF1 induces host cell vulnerability through synthetic lethality.bioRxiv [Preprint]. 2020 Nov 6;221(11):2020.10.12.336487. doi: 10.1101/2020.10.12.336487. bioRxiv. 2020. Update in: J Cell Biol. 2022 Nov 7;221(11):e202011050. doi: 10.1083/jcb.202011050 PMID: 33173868 Free PMC article. Updated. Preprint.
References
-
- Appenzeller-Herzog, C., and H. P. Hauri. 2006. The ER-Golgi intermediate compartment (ERGIC): in search of its identity and function. J. Cell Sci. 119:2173-2183. - PubMed
-
- Dascher, C., and W. E. Balch. 1994. Dominant inhibitory mutants of ARF1 block endoplasmic reticulum to Golgi transport and trigger disassembly of the Golgi apparatus. J. Biol. Chem. 269:1437-1448. - PubMed
-
- Feldmann, H., T. W. Geisbert, P. B. Jahrling, H. D. Klenk, S. V. Netesov, C. J. Peters, A. Shanchez, R. Swanepoel, and V. E. Volchkov. 2004. Filoviridae, p. 645-653. In C. Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball (ed.), Virus taxonomy: VIIIth report of the International Committee on Taxonomy of Viruses. Elsevier/Academic Press, London, United Kingdom.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
