

The midregion, nuclear localization sequence, and C terminus of PTHrP regulate skeletal development, hematopoiesis, and survival in mice
- PMID: 20145205
- PMCID: PMC3140789
- DOI: 10.1096/fj.09-147033
The midregion, nuclear localization sequence, and C terminus of PTHrP regulate skeletal development, hematopoiesis, and survival in mice
Abstract
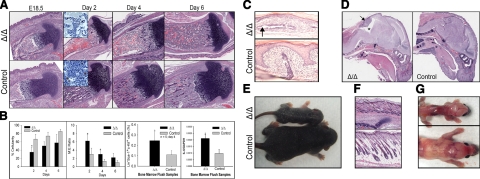
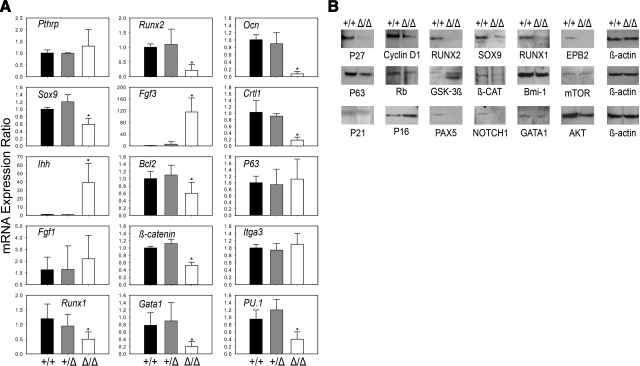
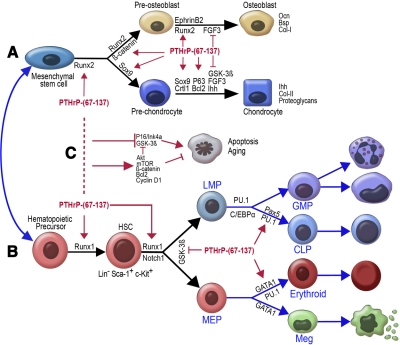
The functions of parathyroid hormone-related protein (PTHrP) on morphogenesis, cell proliferation, apoptosis, and calcium homeostasis have been attributed to its N terminus. Evidence suggests that many of these effects are not mediated by the N terminus but by the midregion, a nuclear localization sequence (NLS), and C terminus of the protein. A knock-in mouse lacking the midregion, NLS, and C terminus of PTHrP (Pthrp(Delta/Delta)) was developed. Pthrp(Delta/Delta) mice had craniofacial dysplasia, chondrodysplasia, and kyphosis, with most mice dying by d 5 of age. In bone, there were fewer chondrocytes and osteoblasts per area, bone mass was decreased, and the marrow was less cellular, with erythroid hypoplasia. Cellular proliferation was impaired, and apoptosis was increased. Runx2, Ocn, Sox9, Crtl1, beta-catenin, Runx1, ephrin B2, cyclin D1, and Gata1 were underexpressed while P16/Ink4a, P21, GSK-3beta, Il-6, Ffg3, and Ihh were overexpressed. Mammary gland development was aberrant, and energy metabolism was deregulated. These results establish that the midregion, NLS, and C terminus of PTHrP are crucial for the commitment of osteogenic and hematopoietic precursors to their lineages, and for survival, and many of the effects of PTHrP on development are not mediated by its N terminus. The down-regulation of Runx1, Runx2, and Sox9 indicates that PTHrP is a modulator of transcriptional activation during stem cell commitment.
Figures


Similar articles
-
Severe growth retardation and early lethality in mice lacking the nuclear localization sequence and C-terminus of PTH-related protein.Proc Natl Acad Sci U S A. 2008 Dec 23;105(51):20309-14. doi: 10.1073/pnas.0805690105. Epub 2008 Dec 17. Proc Natl Acad Sci U S A. 2008. PMID: 19091948 Free PMC article.
-
Absence of PTHrP nuclear localization and C-terminus sequences leads to abnormal development of T cells.Biochimie. 2017 Jul;138:13-19. doi: 10.1016/j.biochi.2017.04.005. Epub 2017 Apr 10. Biochimie. 2017. PMID: 28408247
-
DNA damage checkpoint pathway modulates the regulation of skeletal growth and osteoblastic bone formation by parathyroid hormone-related peptide.Int J Biol Sci. 2018 Apr 5;14(5):508-517. doi: 10.7150/ijbs.23318. eCollection 2018. Int J Biol Sci. 2018. PMID: 29805302 Free PMC article.
-
PTHrP and skeletal development.Ann N Y Acad Sci. 2006 Apr;1068:1-13. doi: 10.1196/annals.1346.002. Ann N Y Acad Sci. 2006. PMID: 16831900 Review.
-
The biological action of parathyroid hormone-related peptide (PTHrP) and fibroblast growth factor receptor 3 (FGFR3) on bone and cartilage.Kaibogaku Zasshi. 2000 Oct;75(5):415-25. Kaibogaku Zasshi. 2000. PMID: 11155687 Review.
Cited by
-
Single-cell RNA sequencing of human non-hematopoietic bone marrow cells reveals a unique set of inter-species conserved biomarkers for native mesenchymal stromal cells.Stem Cell Res Ther. 2023 Aug 30;14(1):229. doi: 10.1186/s13287-023-03437-x. Stem Cell Res Ther. 2023. PMID: 37649081 Free PMC article.
-
Parathyroid Hormone-Related Peptide-Linked Hypercalcemia in a Melanoma Patient Treated With Ipilimumab: Hormone Source and Clinical and Metabolic Correlates.Semin Oncol. 2015 Dec;42(6):909-14. doi: 10.1053/j.seminoncol.2015.09.006. Epub 2015 Sep 8. Semin Oncol. 2015. PMID: 26615135 Free PMC article.
-
PTHrP intracrine actions divergently influence breast cancer growth through p27 and LIFR.Breast Cancer Res. 2024 Feb 26;26(1):34. doi: 10.1186/s13058-024-01791-z. Breast Cancer Res. 2024. PMID: 38409028 Free PMC article.
-
PTHrP induces STAT5 activation, secretory differentiation and accelerates mammary tumor development.Breast Cancer Res. 2022 Apr 19;24(1):30. doi: 10.1186/s13058-022-01523-1. Breast Cancer Res. 2022. PMID: 35440032 Free PMC article.
-
Phospholipase A2-activating protein is associated with a novel form of leukoencephalopathy.Brain. 2017 Feb;140(2):370-386. doi: 10.1093/brain/aww295. Epub 2016 Dec 21. Brain. 2017. PMID: 28007986 Free PMC article.
References
-
- Suva L. J., Winslow G. A., Wettenhall R. E., Hammonds R. G., Moseley J. M., Diefenbach-Jagger H., Rodda C. P., Kemp B. E., Rodriguez H., Chen E. Y. A parathyroid hormone-related protein implicated in malignant hypercalcemia: cloning and expression. Science. 1987;237:893–896. - PubMed
-
- Martin T. J., Ebeling P. R., Rodda C. P., Kemp B. E. Humoral hypercalcemia of malignancy: involvement of a novel hormone. Aust N Z J Med. 1988;18:287–295. - PubMed
-
- Wysolmerski J. J., Stewart A. F. The physiology of parathyroid hormone-related protein: an emerging role as a developmental factor. Annu Rev Physiol. 1998;60:431–460. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
