

Conserved and divergent features of the structure and function of La and La-related proteins (LARPs)
- PMID: 20138158
- PMCID: PMC2860065
- DOI: 10.1016/j.bbagrm.2010.01.011
Conserved and divergent features of the structure and function of La and La-related proteins (LARPs)
Abstract
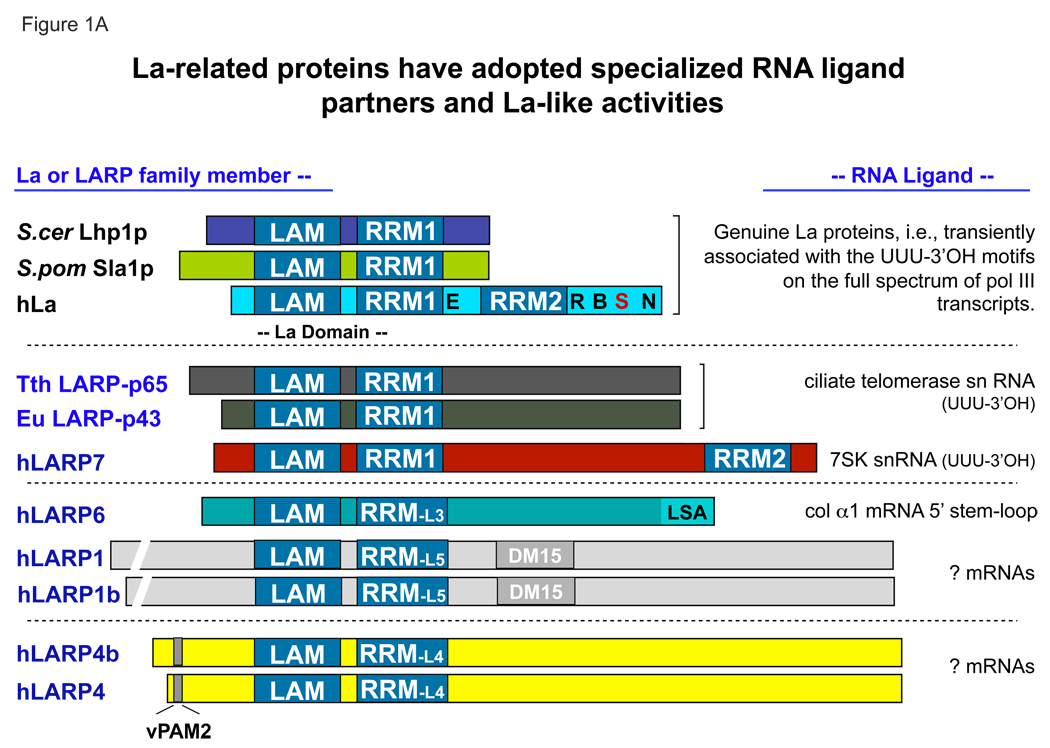
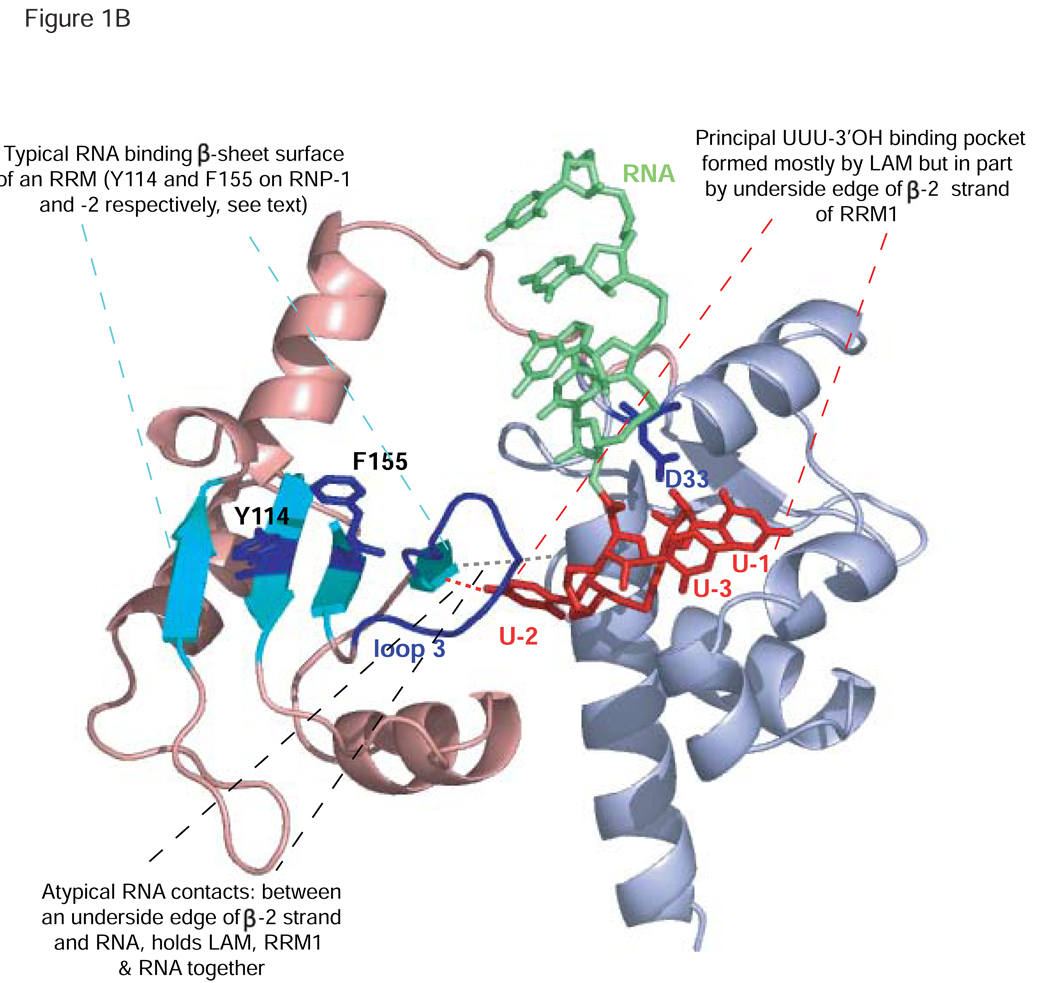
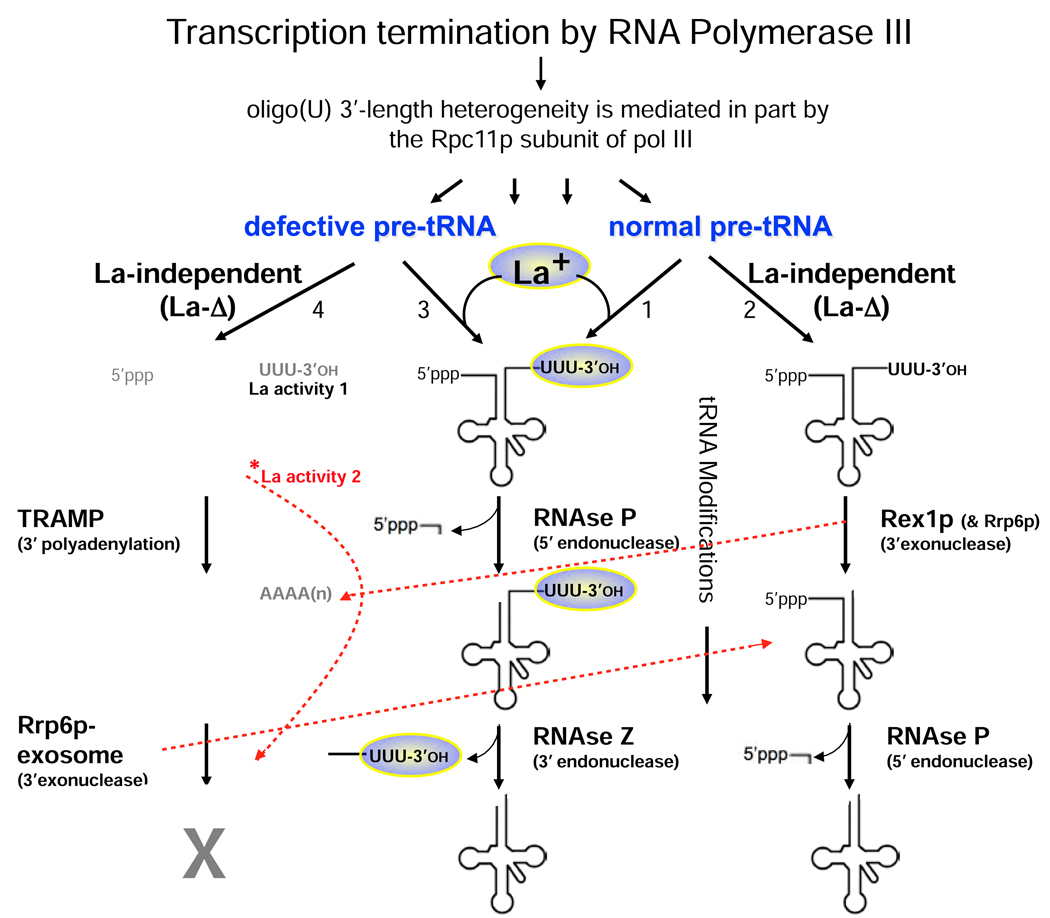
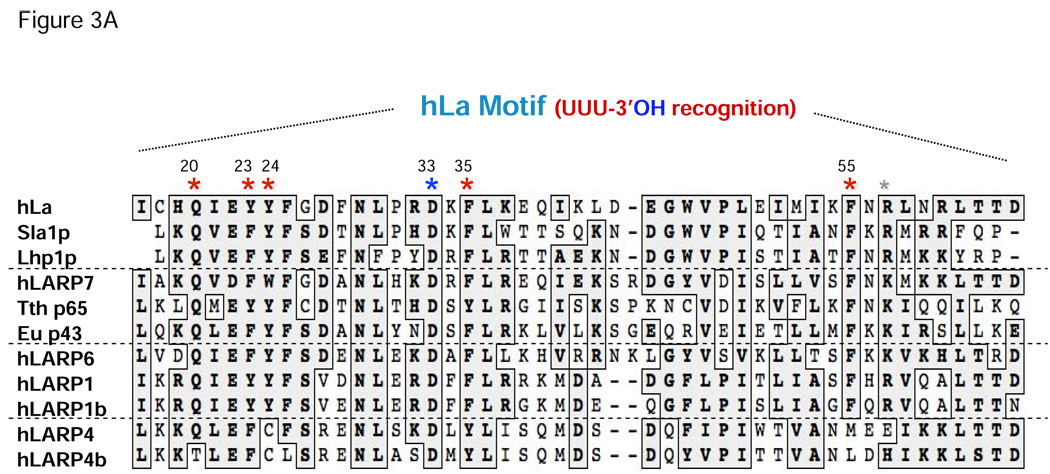
Genuine La proteins contain two RNA binding motifs, a La motif (LAM) followed by a RNA recognition motif (RRM), arranged in a unique way to bind RNA. These proteins interact with an extensive variety of cellular RNAs and exhibit activities in two broad categories: i) to promote the metabolism of nascent pol III transcripts, including precursor-tRNAs, by binding to their common, UUU-3'OH containing ends, and ii) to modulate the translation of certain mRNAs involving an unknown binding mechanism. Characterization of several La-RNA crystal structures as well as biochemical studies reveal insight into their unique two-motif domain architecture and how the LAM recognizes UUU-3'OH while the RRM binds other parts of a pre-tRNA. Recent studies of members of distinct families of conserved La-related proteins (LARPs) indicate that some of these harbor activity related to genuine La proteins, suggesting that their UUU-3'OH binding mode has been appropriated for the assembly and regulation of a specific snRNP (e.g., 7SK snRNP assembly by hLARP7/PIP7S). Analyses of other LARP family members suggest more diverged RNA binding modes and specialization for cytoplasmic mRNA-related functions. Thus it appears that while genuine La proteins exhibit broad general involvement in both snRNA-related and mRNA-related functions, different LARP families may have evolved specialized activities in either snRNA or mRNA-related functions. In this review, we summarize recent progress that has led to greater understanding of the structure and function of La proteins and their roles in tRNA processing and RNP assembly dynamics, as well as progress on the different LARPs.
Figures
Similar articles
-
La proteins couple use of sequence-specific and non-specific binding modes to engage RNA substrates.RNA Biol. 2021 Feb;18(2):168-177. doi: 10.1080/15476286.2019.1582955. Epub 2019 Mar 18. RNA Biol. 2021. PMID: 30777481 Free PMC article. Review.
-
Conservation of RNA chaperone activity of the human La-related proteins 4, 6 and 7.Nucleic Acids Res. 2013 Oct;41(18):8715-25. doi: 10.1093/nar/gkt649. Epub 2013 Jul 25. Nucleic Acids Res. 2013. PMID: 23887937 Free PMC article.
-
Precursor-product discrimination by La protein during tRNA metabolism.Nat Struct Mol Biol. 2009 Apr;16(4):430-7. doi: 10.1038/nsmb.1573. Epub 2009 Mar 15. Nat Struct Mol Biol. 2009. PMID: 19287396 Free PMC article.
-
A comprehensive analysis of the La-motif protein superfamily.RNA. 2009 May;15(5):750-64. doi: 10.1261/rna.1478709. Epub 2009 Mar 19. RNA. 2009. PMID: 19299548 Free PMC article.
-
The La and related RNA-binding proteins (LARPs): structures, functions, and evolving perspectives.Wiley Interdiscip Rev RNA. 2017 Nov;8(6):10.1002/wrna.1430. doi: 10.1002/wrna.1430. Epub 2017 Aug 7. Wiley Interdiscip Rev RNA. 2017. PMID: 28782243 Free PMC article. Review.
Cited by
-
Hydrogen peroxide induces La cytoplasmic shuttling and increases hepatitis C virus internal ribosome entry site-dependent translation.J Gen Virol. 2016 Sep;97(9):2301-2315. doi: 10.1099/jgv.0.000556. Epub 2016 Jul 18. J Gen Virol. 2016. PMID: 27436793 Free PMC article.
-
Nuclear noncoding RNA surveillance: is the end in sight?Trends Genet. 2012 Jul;28(7):306-13. doi: 10.1016/j.tig.2012.03.005. Epub 2012 Apr 2. Trends Genet. 2012. PMID: 22475369 Free PMC article. Review.
-
xRRM: a new class of RRM found in the telomerase La family protein p65.RNA Biol. 2013 Mar;10(3):353-9. doi: 10.4161/rna.23608. Epub 2013 Jan 17. RNA Biol. 2013. PMID: 23328630 Free PMC article. Review.
-
The yeast La related protein Slf1p is a key activator of translation during the oxidative stress response.PLoS Genet. 2015 Jan 8;11(1):e1004903. doi: 10.1371/journal.pgen.1004903. eCollection 2015 Jan. PLoS Genet. 2015. PMID: 25569619 Free PMC article.
-
Viral-host interactions that control HIV-1 transcriptional elongation.Chem Rev. 2013 Nov 13;113(11):8567-82. doi: 10.1021/cr400120z. Epub 2013 Jun 24. Chem Rev. 2013. PMID: 23795863 Free PMC article. Review.
References
-
- Mattioli M, Reichlin M. Heterogeneity of RNA protein antigens reactive with sera of patients with systemic lupus erythematosus. Description of a cytoplasmic nonribosomal antigen. Arthritis Rheum. 1974;17:421–429. - PubMed
-
- Romero V, Fellows E, Jenne DE, Andrade F. Cleavage of La protein by granzyme H induces cytoplasmic translocation and interferes with La-mediated HCV-IRES translational activity. Cell Death Differ. 2009;16:340–348. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
